4190531
Description
Flashcards by Mitra Shadloo, updated more than 1 year ago
|
|
Created by Mitra Shadloo
over 8 years ago
|
|
| Question | Answer |
| Proteine: Bedeutung und Funktionen | "Arbeitsmoleküle" aller lebenden Zellen. Funktionen sind: Katalyse (Enzyme), Strukturgebung (Bindegewebe), Informationsübertragung (Signalkaskaden), Immunabwehr (Antikörper), Muskelkontraktion (Aktin/Myosin), Stofftransport (Hämoglobin, Ionenkanal) |
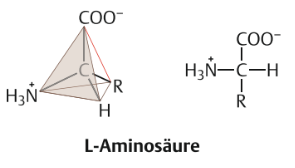
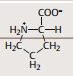
| Aminosäuren Definition und Aufbau | Verbindungen bezeichnet, die ein C-Atom enthalten, das umgeben ist von einer Aminogruppe (-NH2), Carboxylgruppe (-COOH), H-Atom & Rest R, der für die jeweilige Aminosäure charakteristisch ist. |
| Aminosäuren Nomenklatur | Man bezeichnet dieses Kohlenstoffatom, da es in der Strukturformel auf die Carboxylgruppe folgt, als α-C-Atom und die daran gebundenen Gruppen als α-Aminogruppe oder α-Carboxylgruppe |
| Die Aminogruppe Eigenschaften | Die Aminogruppe exponiert ein freies Elektronenpaar, mitunter als seitlicher Strich am Stickstoffatom symbolisiert. Dieses Elektronenpaar kann leicht ein Proton, H+, aufnehmen. Die Aminogruppe erhält dadurch eine positive Ladung und wird zur NH3+-Gruppe. |
| Die Carboxylgruppe Eigenschaften | Ist für den sauren Charakter der AS verantwortlich, gibt leicht ein Proton ab, wobei an der Carboxylgruppe ein überzähliges Elektron & damit eine negative Ladung zurückbleibt. AS sind somit in der Lage, Protonen sowohl aufzunehmen als auch abzugeben |
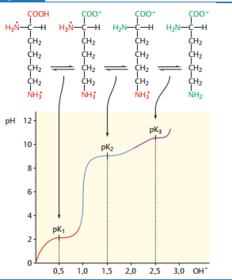
| pK-Wert | Jede Amino- & jede Carboxylgruppe ist bei einem bestimmten, für die jeweilige Gruppe charakteristischen pH-Wert genau zur Hälfte protoniert, die andere Hälfte der Moleküle hat das Proton abgegeben. Diesen pH-Wert = pK-Wert der chemischen Gruppe. |
| Ladung der AS in Bezug auf den pH-Wert | Aminosäuren sind bei niedrigem pH positiv geladen, weil die Amino- und die Carboxylgruppe protoniert sind. Bei steigendem pH-Wert werden die Protonen schrittweise abgegeben |
| AS als Puffer | Lösungen von Aminosäuren haben in der Nähe ihrer pK-Werte eine optimale Pufferkapazität. In größerer Entfernung der jeweiligen pK-Werte puffern Lösungen von Aminosäuren hingegen nicht. |
| Mehrere pK-Werte | Aminosäuren mit mehreren Amino- oder Carboxylgruppen (z. B. Glutaminsäure, Histidin, Lysin) haben mehr als zwei pK-Werte (Histidin z. B. 3). |
| Titrationskurve Lysin | |
| Isoelektrischer Punkt | Den pH-Wert, bei dem eine Aminosäure als Zwitterion vorliegt, die Nettoladung also Null ist, bezeichnet man als isoelektrischen Punkt. |
| Bei physiologischem pH-Wert... | Bei physiologischen pH-Werten (pH 7,2 – 7,4) liegen Aminosäuren als Zwitterionen vor |
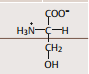
| Allgemeine Strukturformel einer L-Aminosäure | |
| Proteinogene AS | Als proteinogene Aminosäuren werden die 21 Aminosäuren bezeichnet, die bei der Proteinbiosynthese als Proteinbausteine zum Einsatz kommen |
| Ungeladene (neutrale) Aminosäuren | Innerhalb dieser Gruppe lassen sich unpolare und polare Aminosäuren unterscheiden |
| Aliphatische AS Eigenschaften Wer gehört dazu? | Aliphatische Aminosäuren sind unpolar und reaktionsträge, weil ihre Reste keine reaktiven Strukturen aufweisen. Glycin, Alanin, Valin, Leucin, Isoleucin |
| Glycin | Glycin = einzige nicht chirale AS, denn R = H. |
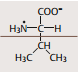
| Alanin | Alanin: R = CH3 |
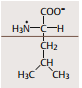
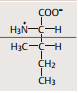
| Valin | ausgesprochen hydrophob. Teil H2O-abweisender (z. B. membrandurch-spannender) Proteinsegmente |
| Leucin (Eigenschaften siehe Valin) | |
| Isoleucin (Eigenschaften siehe Valin) | |
| Aromatische Aminosäuren Eigenschaften Wer gehört dazu? | Tyrosin, Phenylalanin und Tryptophan enthalten jeweils ein aromatisches Ringsystem, also ein ebenes Ringsystem mit delokalisierten π-Elektronen. |
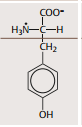
| Tyrosin | Kann aus Phenylalanin gebildet werden. polar, aber ungeladen, da OH-gruppe bei physio. pH (ca. 7,4) nicht ionisiert. |
| Tyrosin weitere Eigenschaften | OH-gruppe kann reversibel eine Phosphatgruppe aufnehmen.. Aktivität vieler Proteine reguliert, indem bestimmte Tyrosine phosphoryliert werden. Aus Tyrosin werden die Schilddrüsenhormone und Katecholamine gebildet. Viele wichtige Pharmaka sind Derivate des Tyrosins. |
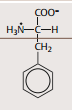
| Phenylalanin | unpolar |
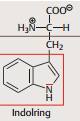
| Tryptophan | unpolar |
| Amide | Asparagin und Glutamin sind Amide der geladenen Aminosäuren Asparaginsäure und Glutaminsäure. Die Amide sind zwar polar, tragen an ihren Resten aber keine Ladung. |
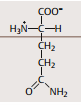
| Asparagin | in vielen Proteinen = Verbindungsstelle -> CHseitenketten |
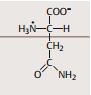
| Glutamin | wichtigste Transportform von Stickstoff |
| Hydroxylierte Aminosäuren | Serin & Threonin + aromatische AS Tyrosin. OH-Gruppe von Serin & Threonin kann Kohlenhydrate binden + reversibel phosphoryliert werden. Viele regulatorisch wichtige Proteine werden durch PH-lierung bestimmter Serine oder Threonine an- oder abgeschaltet. |
| Serin | |
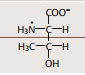
| Threonin | |
| Schwefelhaltige Aminosäuren | Methionin, Cystein & Prolin. Über S-Atom sind beide in der Lage, sich innerhalb von Proteinen an der Bindung von Metallionen zu beteiligen |
| Cystein Eigenschaften | trägt mit seiner SH-Gruppe (= Sulfhydryl-Gruppe) wesentlich zur Stabilität einer Reihe von extrazellulären Proteinen bei. Unter oxidierenden Bedingungen können sich zwei Cysteine unter Ausbildung einer Disulfidbrücke zusammenlagern. Unter reduzierenden Bedingungen -> wieder SH-Gruppen |
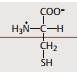
| Cystein Strukturformel | |
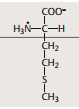
| Methionin | Beteiligt an der Methylierung wichtiger Moleküle |
| Prolin Eigenschaften | Stickstoff in Ringsystem eingebunden, keine freie Aminogruppe mehr. Innerhalb von Proteinen: oft an Stellen, wo AS-Kette einen Knick bildet |
| Prolin Strukturformel | |
| Geladene Aminosäuren | Hierzu zählen die drei basischen Aminosäuren Lysin, Arginin und Histidin sowie die beiden sauren Aminosäuren Asparaginsäure und Glutaminsäure |
| Basische AS Eigenschaften Wer gehört dazu? | Lysin, Arginin und Histidin enthalten in ihren Seitenketten Stickstoffatome, die ein freies Elektronenpaar exponieren. -> leicht ein Proton binden & positive Ladung aufnehmen -> haben einen basischen Charakter. |
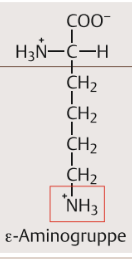
| Lysin Eigenschaften | Exponiert mit seiner Seitenkette eine primäre Aminogruppe. Diese ist mit dem ε-C-Atom des Lysins verbunden und wird deshalb als ε-Aminogruppe bezeichnet. Bei physiologischem pH positiv geladen. |
| Lysin Strukturformel | |
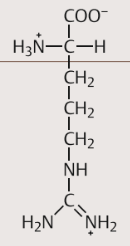
| Arginin Eigenschaften 1 | Bei physiologischen pH positiv geladen. als freie Aminosäure von besonderem biochemischen Interesse: Durch hydro-lytische Abspaltung der stickstoffhaltigen Gruppe (der GuanidinoGruppe) wird in der Leber aus Arginin der Harnstoff gebildet |
| Arginin Eigenschaften 2 | + in den Endothelien der Blutgefäße Ausgangsstoff für Synthese von (NO). Dieses relaxiert die benachbarten glatten Gefäßmuskelzellen und löst so eine Weitstellung des Gefäßes aus. Dadurch spielt NO eine bedeutende Rolle in der Regulation des Blutdrucks. |
| Arginin Strukturformel | |
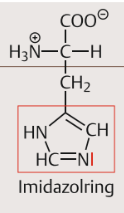
| Histidin Eigenschaften | Der Imidazolring des Histidins kann bei physiologischen pH-Werten leicht Protonen aufnehmen und wieder abgeben. In vielen Proteinen ist er an der Bindung von Metallionen beteiligt. bei physiologischen pH-Werten nur teilweise protoniert |
| Histidin Eigenschaften 2 | in Enzymen oft an Stellen, an denen die gezielte Übertragung eines Protons erforderlich ist. im Gegensatz zu den anderen aromatischen Aminosäuren ist Histidin aber ausgesprochen hydrophil. |
| Histidin Strukturformel | |
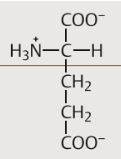
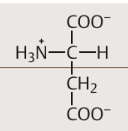
| Saure Aminosäuren Wer gehört dazu? Eigenschaften | Asparaginsäure und Glutaminsäure. enthalten eine Carboxylgruppe, deren Proton leicht abdissoziiert. Dabei entsteht Aspartat bzw. Glutamat. Aufgrund ihres Gehalts an Aspartat und Glutamat sind die meisten Proteine negativ geladen |
| Glutamat Strukturformel | |
| Aspartat Strukturformel | |
| Wie entstehen nicht essenzielle AS? | z. B. Alanin, entstehen durch Transaminierung. Andere proteinogene Aminosäuren werden im Metabolismus des Menschen in z. T. recht komplizierten Stoffwechselwegen gebildet, z. B. Prolin. |
| Essenzielle AS | können im Stoffwechsel des Menschen unter keinen Umständen synthetisiert werden. Sie müssen deshalb unbedingt in hinreichender Menge mit der Nahrung aufgenommen werden. |
| Unbedingt essenziell | Valin, Leucin, Isoleucin, Phenylalanin, Tryptophan, Methionin, Threonin, Lysin |
| bedingt essenziell bei Fehlen von Phenylalanin bzw. Methionin | Tyrosin (kann nicht mehr durch Hydroxylierung aus Phenylalanin entstehen) & Cystein (kann nur gebildet werden, sofern Methionin in ausreichenden Mengen zur Verfügung steht) |
| bei weitgehendem oder völligem Fehlen in der Nahrung und bei Säuglingen | Histidin & Arginin (können dann nicht in ausreichender Menge synthetisiert werden, wenn sie in der Nahrung vollständig oder weitgehend fehlen. Sie sind für Säuglinge essenziell) |
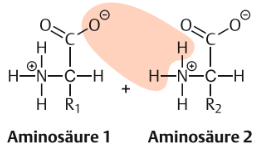
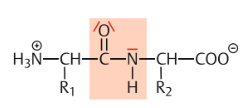
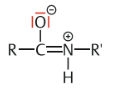
| Peptidbindungen Wo werden sie gebildet? Was passiert bei der Reaktion/welche Gruppen reagieren? | Peptidbindungen werden in den Zellen normalerweise unter Vermittlung von Ribosomen gebildet. Entspricht einer Verbindung zwischen der Carboxylgruppe einer Aminosäure und der Aminogruppe einer anderen Aminosäure unter Abspaltung von Wasser |
| Peptidbindung Edukte | |
| Peptidbindung Produkte | |
| Partielle Doppelbindung | Das freie Elektronenpaar des Stickstoffs wird durch die hohe Elekttronegativität des Sauerstoffs in die Bindung zwischen N- und C-Atom hineingezogen, wodurch die C ═ O und die N-H-Gruppe ihre freie Drehbarkeit gegeneinander verlieren. |
| Wie sieht die Peptidbindung jetzt aus? (Bild) | |
| Was für eine Konsequenz hat dieser partielle Doppelbindungscharakter? | Peptidbindungen sind starre, ebene Strukturelemente. Dadurch haben alle Peptide und Proteine auch nur begrenzte Möglichkeiten, sich im Raum zu definierten Strukturen anzuordnen. |
| Was für eine Rolle spielen Wasserstoffbrücken? | Das O- & O-Atom der Peptidbindungen -> Bildung von Wasserstoffbrücken beteiligt, die gegenüberliegende Peptid-bindungen verbinden und zur Bildung definierter Proteinstukturen führen können. Diese sind aber nicht sehr stabil und brechen leicht bei Hitze auf -> Denaturierung! |
| Primärstruktur | Unter der Primärstruktur eines Proteins versteht man seine Aminosäuresequenz. reicht nicht für die Erhaltung der Proteinfunktion aus! Hierfür ist die Sekundär-, Tertiär- und Quartärstruktur erforderlich. Bei Denaturierung bleibt Primärstruktur erhalten. Funktion ist weg |
| Sekundärstruktur | die regelmäßigen Strukturen innerhalb von Polypeptiden, die sich aufgrund von WSB zwischen Peptidbindungen ausbilden. Die verschiedenen Abschnitte einer Aminosäuresequenz zeigen in der Regel unterschiedliche Sekundärstrukturen |
| Rolle der AS-Reste bei Ausbildung der Sekundärstruktur | können die Ausbildung einer bestimmten Sekundärstruktur zwar wesentlich begünstigen, an der Ausbildung der entscheidenden Wasserstoffbrücken der Sekundärstrukturen sind sie aber nicht beteiligt. |
| Sekundärstrukturelemente. | α-Helix, β-Faltblatt (engl. β-sheet) U-förmige Verbindungsstücke (engl. loop bzw. turn), im Deutschen meist als Schleife bezeichnet. |
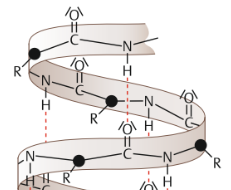
| α-Helix | schraubig gewundene Abschnitte einer ASkette. jeweils 3,6 AS eine Windung. Alle Seitenketten der beteiligten Aminosäuren weisen nach außen. ausschließlich durch die Wasserstoffbrücken stabilisiert, die sich zwischen den Peptidbindungen der einzelnen Windungen ausbilden |
| WSB und Reste bei einer α-Helix | Die Wasserstoffbrücken bilden sich also bei einer α-Helix innerhalb einer ASkette aus. Die ASreste stehen hingegen für Wechselwirkungen mit anderen ASketten zur Verfügung. |
| α-Helix | |
| Ausnahme Prolin | Prolin ist die einzige Aminosäure, deren Peptidbindung kein Wasserstoffatom aufweist und die sich deshalb auch nicht an der Bildung einer Wasserstoffbrücke beteiligen kann. Aus diesem Grund kann bereits ein einzelnes Prolin eine α-Helix unterbrechen: Es ist ein „Helixbrecher“. |
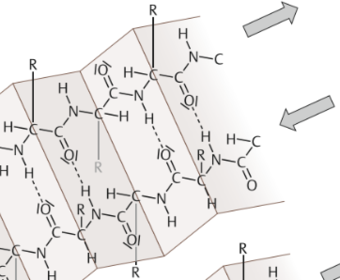
| β-Faltblatt | ASketten lagern sich in weitgehend gestreckter Konformation nebeneinander. durch WSB stabilisiert, die sich zwischen zwei parallel oder auch antiparallel liegenden ASketten ausbilden. Die Aminosäurereste ragen dann abwechselnd nach oben und nach unten |
| β-Faltblatt Struktur Bild | |
| Schleifen | Schleifen sind U-förmige Abschnitte der Aminosäurekette, die die α-Helices und β-Faltblattstrukturen eines Proteins miteinander verbinden. Einfachster Fall: β-Schleife, besteht aus 4 AS, wobei die erste und die vierte Aminosäure durch eine Wasserstoffbrücke verbunden sind |
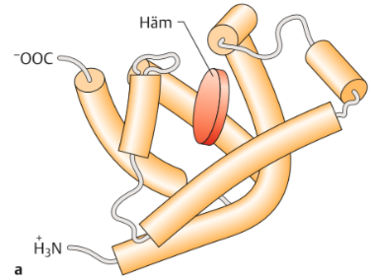
| Tertiärstruktur | beschreibt die räumliche Struktur einer kompletten ASkette, einschließlich der Anordnung sämtlicher ASreste. wird durch Wechselwirkungen der Aminosäurereste stabilisiert |
| Welche Welchselwirkungen kann es geben? | hydrophobe Wechselwirkungen Disulfidbrücken ionische Wechselwirkungen van der Waals-Kräfte |
| Bsp: Tertiärstruktur eines Hämboglobins | |
| Hydrophobe Wechselwirkungen | In der Regel zeigen die hydrophoben Aminosäurereste ins Innere des Proteins, die polaren oder geladenen Aminosäurereste nach außen. Der Grund für diese Anordnung der Aminosäurereste sind Wechselwirkungen mit den umgebenden Wassermolekülen. |
| Rolle hydrophober Wechselwirkungen | verantwortlich für die Zusammenlagerung hydrophober Moleküle oder hydrophober Teile von Molekülen in einer wässrigen Umgebung. In vielen Fällen liefern die hydrophoben Wechselwirkungen den größten Beitrag zur Stabilität von Proteinen. |
| Disulfidbrücke und ionische Wechselwirkungen | Disulfidbrücken zwischen zwei Cysteinen (kovalente Bindung) verbinden oft die ASketten extrazellulärer Proteine, z. B. im Insulin und in Antikörpern. Ionische Wechselwirkungen (stark!) aufgrund gegensätzlicher Ladung treten bei nahezu allen Proteinen auf. |
| Van-der-Waals-Kräfte | vergleichsweise schwach. Nachdem sich die hydrophoben Anteile innerhalb eines Proteinmoleküls durch hydrophobe Wechselwirkungen zusammen gelagert haben, tragen sie dazu bei, die genaue räumliche Anordnung der Atomgruppen festzulegen. |
| Wie werden sie hervorgerufen? | Van-der-Waals-Kräfte werden durch minimale Ladungsinhomogenitäten innerhalb der Atome hervorgerufen. |
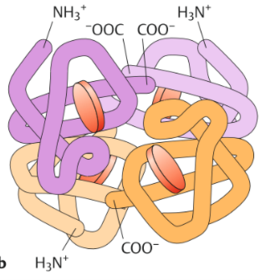
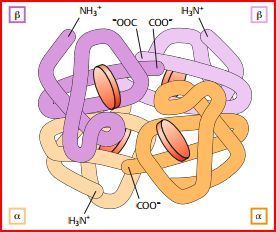
| Quartärstruktur | beschreibt die Zahl und die Anordnung der verschiedenen ASketten in einem Proteinkomplex. ist also nur gegeben, wenn sich mehrere Aminosäureketten zu einem Komplex zusammenlagern. |
| Quartärstruktur Hämoglobin | |
| Co- und Posttranslationale Modifikationen Definition | Enzymatisch katalysierte Veränderungen der Proteine während oder nach der Translation nennt man co- oder posttranslationale Modifikationen. Viele Proteine müssen während oder nach der Translation modifiziert werden, um funktionsfähig zu werden |
| Posttranslational Proteolytische Prozessierung und limitierte Proteolyse: Methionin | Alle Proteine enthalten unmittelbar nach der Translation ein aminoterminales Methionin aufgrund der Verwendung des Codons AUG als Translationsstart. Diese Aminosäure wird bei den meisten Proteinen durch eine Methionin-spezifische Aminopeptidase entfernt. |
| Posttranslational Proteolytische Prozessierung und limitierte Proteolyse: Peptidhormone | Bildung von Peptidhormonen (z. B. Insulin) aus Vorläufermolekülen durch Proteolyse. Vom Präproinsulin wird beim cotranslationalen Transport in das ER die Signalsequenz entfernt, wodurch das Proinsulin entsteht. Weitere Schnitte: C-Kette ab -> fertiges Insulin |
| Posttranslational Proteolytische Prozessierung und limitierte Proteolyse: Inaktive Enzyme (Zymogene) | Aktivierung von Protease-Vorstufen zu Proteasen durch Proteolyse: Verdauungspeptidasen (z. B. Trypsinogen → Trypsin), Blutgerinnungskaskade, intrazellulär spielt dies bei der Auslösung der Apoptose durch Caspasen eine Rolle |
| Posttranslational Hydroxylierung: | Lysin- und Prolinreste des Kollagens werden durch α-Ketoglutaratabhängige Dioxygenasen hydroxyliert, die durch Vitamin C funktionsfähig gehalten werden. Nur so kann die Tripelhelix des Kollagens durch WSB stabilisiert werden |
| Posttranslational Carboxylierung: | Vit.-K-abhängige Carboxylierung von Glutamat-R ist für die Funktion einiger Calcium-bindender Proteine (z. B. die Blutgerinnungsfaktoren IX, X, VII, Prothrombin, Proteine C/S) essenziell. entstandene γ-Carboxyglutamat kann Ca2+ gut chelatartig binden -> erhält Funktion |
| Vit. K Antagonisten | Der therapeutische Einsatz von Vitamin-K-Antagonisten führt zu einer Minderung der Blutgerinnungsneigung und dadurch zu einer Verringerung des Thromboserisikos |
| Phosphorylierung: | reversible Phosphorylierung der Aminosäuren Serin, Threonin und Tyrosin zählt zu den wichtigsten Mechanismen zur Regulation der Enzymaktivität. |
| Glykosylierung | Kohlenhydratreste können im ER oder im Golgi-Apparat an die Aminosäuren Serin oder Threonin (O-glykosidisch) oder an Asparagin (N-glykosidisch) geknüpft werden |
| Abbau von Proteinen | Aminosäuren = essenzielle Quelle organischer Stickstoffverbindungen. Proteine sind die wichtigste Quelle für die 8 essenziellen Aminosäuren. |
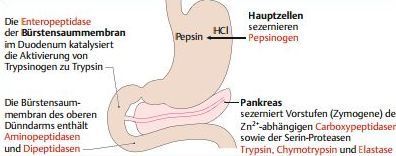
| Hydrolyse der Proteine durch Proteasen | Im Magen: durch Magensäure denaturiert. Dann Hydrolyse durch Pepsin (Endopeptidase), innerhalb der AS-Kette. Im Duodenum: Polypeptide werden hydrolysiert durch Pankreas-Proteasen. |
| Welche Pankreas-Proteasen gibt es? A | A) Serin-Proteasen: 3 Endopeptidasen - Trypsin (aktiviert durch Peptidase auf Bürstensaum der Enterozyten). Chrymotrypsin (aktiviert durch Trypsin). Elastase (hydrolysiert u. a. Protein Elastin, das im Bindegewebe elastische Fasern bildet) |
| Welche Pankreas-Proteasen gibt es? B | Carboxypeptidasen: Carboxypeptidasen A und B sind Exopeptidasen. Sie spalten von ihren Substraten jeweils die carboxyterminale Aminosäure ab. Im Reaktionsmechanismus spielt ein Zink-Ion eine entscheidende Rolle. |
| Was sind die Hydrolyseprodukte der Serin- & Carboxypeptidasen? | überwiegend Oligopeptide, teilweise freie AS. die letzten Hydrolyseschritte laufen an Enterozytenmembran ab. Bürstensaum enthält mehrere Peptidasen, bei denen es sich überwiegend um Aminopeptidasen und um Dipeptidasen handelt |
| Bild | |
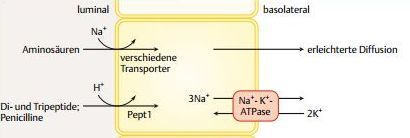
| Resorption der Hydrolyseprodukte Freie AS | Die von den Proteasen und Peptidasen freigesetzten Aminosäuren werden wie die Monosaccharide sekundär-aktiv in einem Symport mit Na+-Ionen transportiert. Indirekt ist der Prozess von der Na+-K+-ATPase der basolateralen Membran abhängig |
| Bild | |
| Resorption der Hydrolyseprodukte Di- & Tripeptide | Der Oligopeptid-Translokator, Pept1, vermittelt einen H+-Symport von Di- und Tripeptiden. Der Transporter akzeptiert Peptide unterschiedlicher AS-Zusammensetzung und arbeitet sehr effizient. |
| Die Struktur des Hämoglobins | Ein Tetramer. Die häufigste Form, HbA1, besteht aus 2 α- & 2 β-Untereinheiten. Jede Untereinheit kann ein O2-Molekül binden. In den Bindestellen lagert sich O2 jeweils an ein zweiwertiges Eisen-Ion (Fe2+) an, das sich im Zentrum eines Porphyrinrings befindet. |
| Bindung von O2 | Dabei wird das zentrale Eisen-Ion nicht oxidiert, sondern bleibt als Fe2+ erhalten. Entsprechend handelt es sich bei der Beladung mit O2 auch nicht um eine Oxidation des Hämoglobins, sondern nur um eine Oxygenierung. |
| Bild | |
| Was ist eine Hämgruppe? | Ein Porphyrinring mit eingelagerten Eisen-Ion wird als Hämgruppe bezeichnet. Hämgruppen sind als prosthetische Gruppen nicht nur im Hämoglobin, sondern auch in den Cytochromen der Atmungskette/Cytochrom P-450 des endoplasmatischen Retikulums enthalten. |
| Wie werden die Hämgruppen im Hämoglobin gebunden? Was bildet den Globin-Anteil des Moleküls? | die Hämgruppen werden im Hämoglobin lediglich durch nicht kovalente Bindungen in hydrophoben Taschen des Proteins festgehalten. Die Aminosäureketten der Untereinheiten bilden den Globin-Anteil des Hämoglobins. |
| Kohlenmonoxid (CO) | In die O2-Bindestelle kann sich auch Kohlenmonoxid (CO) einlagern. Da die Affinität des CO für die Bindestelle 200- bis 300-mal höher ist als die des O2, können bereits geringe CO-Mengen zu einer erheblichen Vergiftung führen. |
| Symptome & Therapie Kohlenmonoxid-Vergiftung | Kopfschmerzen, Bewusstlosigkeit und Kreislaufkollaps. Die Bindung des CO ist vollständig reversibel. Zur Therapie ist deshalb in vielen Fällen ein rasches Verbringen in CO-freie Luft hinreichend. Lebensgefährlich sind in der Regel erst Hb-CO-Anteile von über 60 %. |
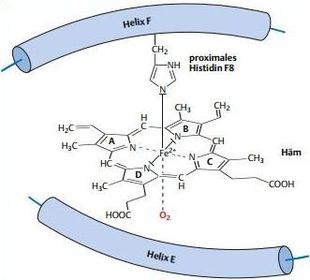
| Struktur der Sauerstoffbindestelle | jede Hämgruppe hat O2 nur an einer Seite des Fe2+ gebunden. Auf der anderen Seite wird das Fe2+ von der Imidazol-gruppe eines Histidins festgehalten. Dieses „proximale“ Histidin gehört zu einer α- Helix (Helix F), das Histidin innerhalb dieser Helix an Position 8 steht, wird es als Histidin F8 bezeichnet. |
| Bild | |
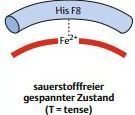
| Auswirkungen der O2-Bindung auf die Ultrastruktur des Hämoglobins T-Zustand | Porphyrinringe sind ebene Strukturen. Im Hämoglobin übt die Imidazolgruppe des Histidins F8 jedoch eine Kraft auf das Fe2+ aus. Die Hämgruppe wölbt sich dem Histidin F8 entgegen. Hämoglobin befindet sich dadurch in einem Spannungszustand, der als T-Zustand bezeichnet wird |
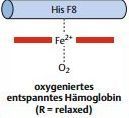
| Was passiert, wenn ein O2 bindet? | Bindet an der anderen Seite der Hämgruppe ein O2, zieht dieses das Fe2+ in die Ebene des Porphyrinrings zurück, und die Hämgruppe ist wieder planar. Dieser Bewegung folgt das Histidin F8, und die Helix F biegt sich auch in Richtung der Hämgruppe. |
| Was für Folgen hat die Konformationsänderung? | Hat weitreichende Verschiebungen im gesamten Hämoglobinmolekül zur Folge, sodass sich letztlich in allen vier UE eine entspanntere Konformation einstellt. R-Zustand. Auch in den O2-freien UE bewegen sich die F-Helices in Richtung der Hämgruppen. Das gesamte Molekül befindet sich daraufhin im R-Zustand. |
| Wie wird die Funktion des Hämoglobins dadurch beeinflusst? | Für die Funktion des Hämoglobins ist es nun von fundamentaler Bedeutung, dass mit dem R-Zustand auch eine erheblich erhöhte O2- Affinität aller vier Untereinheiten verbunden ist. |
| Hämoglobin im O2-freien Zustand | |
| Hämoglobin im O2-gebundenen Zustand | |
| Was ist das besondere an der Wirkung der Konformationsänderung? | Mit Beladung der ersten O2-Bindestelle steigt die Wahrscheinlichkeit, dass das Hämoglobinmolekül den R-Zustand einnimmt. Bei Oxygenierung weiterer Bindestellen nimmt diese nochmals zu. Die O2-Affinität erhöht sich dabei um bis zu drei Größenordnungen. Man spricht von "Kooperativität" |
| Wovon hängt die Menge an O2 ab, die Hämoglobin in der Lunge aufnimmt? | vom Partialdruck des Sauerstoffs in Atemluft bzw. Blut ab, von dem Druck also, unter dem O2 in dem Gasgemisch Luft bzw. Blut steht. O2 diffundiert in Richtung des niedrigeren Drucks, bis die Druckdifferenz ausgeglichen ist. |
| Merke | Das Hämoglobin im Lungenvenenblut ist zu 97 %, d. h. bis nahe an die Grenze seiner Kapazität mit O2 gesättigt. Aufgrund der großen Menge an Hämoglobin, das in den Erythrozyten enthalten ist, kann 1 Liter Blut (1000 ml) bis zu 200 ml (O2) transportieren. |
| In den peripheren Geweben | wird dem Blut O2 entzogen, denn der PO2 beträgt dort nur noch ca. 40 mm Hg. Bei der Abgabe des O2 in den peripheren Geweben gleichen sich die Partialdrücke, wieder nahezu an. Dennoch sinkt die O2-Sättigung im venösen Blut nur auf 75%. |
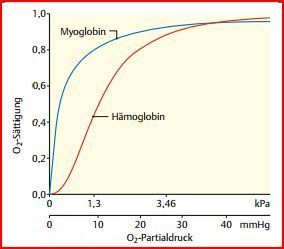
| die O2-Bindungskurve des Hämoglobins... | die die O2-Sättigung (SO2) in Abhängigkeit vom O2-Partialdruck (PO2) darstellt, zeigt einen S-förmigen = sigmoiden Verlauf. |
| Im Bereich der hohen PO2-Werte... Im Bereich der niedrigen PO2-Werte... | ...verläuft die Kurve flach, d. h. die SO2 ändert sich nur geringfügig. ...dagegen verläuft die Kurve steil, d. h. eine geringe Abnahme des PO2 führt zu einer deutlichen Abnahme der SO2. |
| Bei niedrigen PO2- Werten... | wie sie in den peripheren Geweben herrschen, gibt Hämoglobin den gebundenen Sauerstoff also besonders leicht ab. |
| O2-Bindungskurve des Hämoglobins Bild | |
| Eine wesentliche Voraussetzung für den sigmoiden Verlauf der O2-Bindungskurve... | besteht darin, dass jedes Hämoglobinmolekül mehrere Untereinheiten besitzt, die jeweils O2 binden. |
| Die Bindung des ersten O2-Moleküls an eine der UE des Hämoglobins verändert... | die räumliche Struktur sämtlicher UE des Hämoglobins mit der Folge, dass die anderen UE jedes weitere O2- Molekül mit erhöhter Aufnahmebereitschaft binden: Die UE des Hä- moglobins zeigen Kooperativität. |
| Die Kooperativität der Hämoglobin-Untereinheiten ist der Grund dafür, | dass Hämoglobin in einer Umgebung mit sehr niedrigem PO2 zunächst kaum O2 aufnimmt, dass aber bereits mittlere Partialdrücke ausreichen, um Hämoglobin zu einem großen Anteil mit O2 zu beladen |
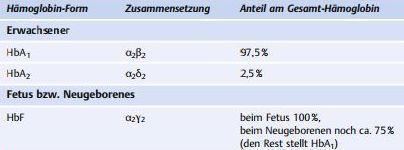
| Die verschiedenen Hämoglobine des Menschen | |
| Das HbF des Fetus... | hat im Vergleich zu HbA1 eine geringere Affinität zu 2,3-BPG und damit eine wesentlich höhere O2-Affinität, was die Umverteilung von O2 vom mütterlichen in den fetalen Kreislauf begünstigt. |
| Myoglobin | kleines Protein , das zu den UE der Hämoglobine große Ähnlichkeiten zeigt. Es enthält auch gleiche Porphyrinringsystem. Allerdings vermittelt es O2-Transport nicht im Blut, sondern in der Herz-/ Skelett-muskulatur. Erleichtert IZ-Transport des O2 zu Mitos. |
| Anders als die Hämoglobine liegt Myoglobin aber stets... | als Monomer vor. Deshalb ist seine O2-Bindungskurve nicht S-förmig wie die der Hämoglobine, sondern eine Hyperbel, O2-Affinität ist deutlich höher als die der Hämoglobine. Kapazität ist aber begrenzt, ermöglicht Aufrechterhaltung eines aeroben Stoffwechsels für wenige Sekunden. |
| Myoglobin kann nicht nur O2 binden, sondern... | auch NO, in einer Reaktion mit Myoglobin-gebundenem O2 wird NO zu Nitrat umgesetzt. Wahrscheinlich verhindert das, dass NO zu den Mitochondrien vordringt und die Atmungskette blockiert. NO ist ein potenter Inaktivator des Komplexes IV der Atmungskette |
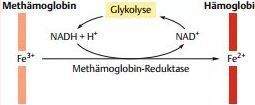
| Schutz des Hämoglobins vor Oxidation | Wenn die Fe2+-Ionen des Hämoglobins zu Fe3+ oxidiert werden, entsteht dunkel gefärbtes Methämoglobin. Methämoglobin kann keinen Sauerstoff binden. |
| Methämoglobin entsteht... | in den Erythrozyten, wenn Hämoglobin-gebundener O2 vom Fe2+-Ion ein Elektron aufnimmt und sich als Superoxidanion (O2– ) ablöst. |
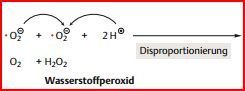
| Das Superoxidanion, O2– , ist.... | ein Sauerstoffradikal (ROS). O2– entsteht auch in anderen Nebenreaktionen des Stoffwechsels, indem ein einzelnes Elektron auf molekularen Sauerstoff übertragen wird. O2– reagiert spontan mit verschiedenen organischen Verbindungen der unmittelbaren Umgebung, schädigt dadurch die Zellen. |
| Superoxidradikal | |
| Wenn zwei Superoxidradikale aufeinander treffen, | reagieren sie sofort unter Bildung eines reduzierten und eines oxidierten Reaktionsprodukts: O2 und H2O2 |
| Wasserstoffperoxid | |
| H2O2 ist ein... | aggressives Oxidationsmittel, das die Zellen ebenfalls erheblich schädigen kann. |
| Ausgehend von Superoxidradikalen kann neben H2O2 auch... | das elektrisch neutrale, aber außerordentlich aggressive Hydroxylradikal (·OH) entstehen |
| Hydroxylradikal | |
| Schutzmechanismen | Die Methämoglobin-Reduktase katalysiert die Reduktion des Fe3+ zu Fe2+. Als Reduktionsmittel dient dabei NADH (das in den Erythrozyten von der Glykolyse bereitgestellt wird). |
| Bild | |
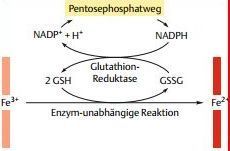
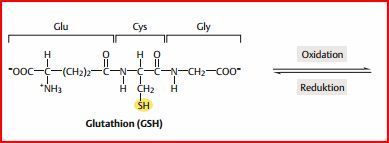
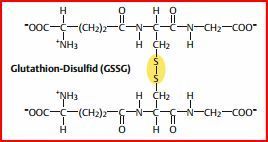
| Glutathion | Trägt auch zum Schutz des Hämoglobins gegen Oxidation, in den Erys enthalten ist. Glutathion ist ein Tripeptid. Kann Methämoglobin reduzieren, indem es direkt mit den Fe-Ionen der Hämgruppen reagiert. Primär dient es dem Schutz der SH-Gruppen des Hämoglobins. |
| Glutathion wird von... | einer Glutathion-Reduktase regeneriert. Das Enzym verwendet dazu NADPH (das aus dem Pentosephosphatweg der Erythrozyten stammt). |
| Bild | |
| Bildung von Heinz-Körpern | wenn der Pentosephosphatweg aufgrund eines Defekts der Glucose-6-phosphat-Dehydrogenase nicht genügend NADPH liefern kann, beeinträchtigt das die Reduktion des Methämoglobins + Schutz der SH-Gruppen des Hämoglobins. Charakteristisch ist Bildung unlöslicher Hämoglobin-Aggregate |
| Wichtige Antioxidanzien sind neben dem Glutathion.... | die Vitamine C (Ascorbinsäure) und E (Tocopherol), sowie das beim Häm-Abbau anfallende Bilirubin. |
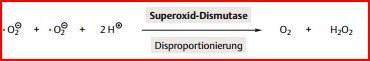
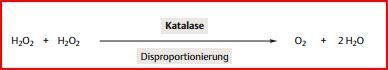
| Zudem ist die Kooperation der Enzyme auch in allen anderen Zellen des Körpers durch... | Superoxid-Dismutase und Katalase gewährleistet. Beide Enzyme katalysieren Disproportionierungen: Die Superoxid-Dismutase katalysiert die Reaktion von zwei Superoxidradikalen (O2-) zu O2 und H2O2. Das H2O2 wird dann mithilfe der Katalase zu O2 und H2O disproportioniert. |
| Superoxid-Dismutase | |
| Katalase | |
| Transport von CO2 | CO2 wird im Blut zu ca. 90 % in Form von Bicarbonat transportiert. Ca. 5 % sind physikalisch gelöst. Nur ca. 5 % des CO2 sind an Hämoglobin gebunden. |
| Bicarbonat entsteht in einer Reaktion... | von CO2 mit Wasser: CO2 + H2O ⇌ HCO3– + H+ |
| Die Reaktion verläuft normalerweise nur sehr langsam. Erythrozyten enthalten aber ein Enzym: | die Carboanhydrase, welches die Reaktion beschleunigt. Eine CA 600 000 Moleküle CO2 zu HCO3-/s umsetzen. Das HCO3– wird von Erys weitgehend an Blut abgegeben. Reaktion ist reversibel. In der Lunge ist CA entscheidend an der Mobilisierung des CO2 aus dem HCO3 – beteiligt. |
| Bicarbonat liegt im Blut in einer Konzentration von ca. 24 mmol/l vor und leistet... | den größten Beitrag zur Stabilisierung des physiologischen pH-Wertes bei 7,4. Zusammen mit dem physikalisch gelösten CO2 repräsentiert es den Bicarbonat-Puffer des Blutes |
| Der zweitwichtigste Faktor für die Aufrechterhaltung des pH-Wertes ist das... | Hämoglobin mit seinem Protonenakzeptor/-donator Histidin der β-Untereinheit. Diesen Beitrag kann Hämoglobin nur leisten, weil es im Blut in außerordentlich hoher Konzentration (160 g/l) vorhanden ist. |
| Transport gebunden an Hämoglobin | Die α-Aminogruppe der jeweils ersten Aminosäure jeder Untereinheit des Hämoglobins kann kovalent ein CO2 binden: Hb-NH2 + CO2 ⇌ Hb-NHCOO– + H+ |
| Die entstandene NHCOO–-Gruppe wird als... | Carbaminogruppe bezeichnet. Nach Bildung der negativ geladenen Carbamatgruppen nimmt das Hämoglobinmolekül bevorzugt die Konformation des T-Zustandes, also die O2-freie Konformation ein. |
| Dieses hat eine bemerkenswerte Konsequenz: | Hämoglobin bindet bevorzugt entweder CO2 oder O2 (Haldane-Effekt). Eine niedrige O2-Konzentration (in O2-armen Geweben) begünstigt die Bindung von CO2. Bei hoher O2-Konzentration (Sättigung des Hämoglobins mit O2 in der Lunge) dagegen ist die Abgabe des CO2 erleichtert. |
| Proteinfaltung Was für eine Funktion hat sie? | Proteinfaltung ist der Prozess, der zur Ausbildung einer funktionellen dreidimensionalen Struktur, d. h. der nativen Struktur, eines Proteins führt. |
| Was ist durch die Gene festgelegt? Was durch die AS-Sequenz? | Durch die Gene ist zunächst einmal nur die Primärstruktur des Proteins, die Aminosäuresequenz, festgelegt. Diese enthält grundsätzlich die gesamte notwendige Information zur Faltung eines Proteins in die funktionelle dreidimensionale Struktur. |
| Aus den bindenden Kräften... | die zwischen bestimmten Abschnitten der Polypeptid-kette bestehen (WSB, Ionen-bindungen, Disulfidbrücken, hydrophobe Wechselwirkungen), ergibt sich eine thermodynamisch stabile, d. h. mit einem Minimum an freier Energie verbundene Konformation=native Struktur des Proteins. |
| Die Faltung erfolgt... Dabei kann es passieren... | ...über eine begrenzte Anzahl geordneter Folgen von Faltungsschritten, bis das Energieminimum des nativen Zustandes erreicht wird. ...dass einige Intermediärprodukte hydro-phobe Bereiche exponieren, die können zur Aggregation einzelner Polypeptidketten führen |
| Folgen von Aggregatbildungen: | Derartige Aggregatbildungen oder Fehlfaltungen beobachtet man bei einer Reihe von neurodegenerativen Erkrankungen, wie z. B. bei Morbus Alzheimer, Morbus Parkinson, Chorea Huntington ect. |
| Wie kann eine Fehlfaltung verhindert werden? | Bei allen Lebewesen haben sich Schutzmechanismen entwickelt, die Fehlfaltungen und gefährliche Aggregatbildungen verhindern sollen. Wichtigste Bestandteile dieses Schutzsystems sind die Chaperone. |
| Das Hauptproblem dabei sind... | ....die hydrophoben AS-Seitenketten. Im richtig gefalteten Protein liegen sie gemeinsam im Inneren, nach außen zum Wasser ist das Protein hydrophil. Wenn eine hydrophobe Aminosäure aus dem Ribosom herauskommt, sucht sie wie ein Öltröpfchen andere hydrophobe Stoffe und lagert sich mit ihnen zusammen. |
| Dies können auch hydrophobe Bereiche... | ...von anderen gerade entstehenden Proteinen sein, zumal die weiteren hydrophoben Aminosäuren des eigenen Proteins noch gar nicht vom Ribosom angehängt wurden. Aggregationen, sind oft irreversibel, die Proteine müssen abge- baut werden, bevor sie jemals ihre aktive Konformation erreicht hatten. |
| Molekulare Chaperone sind... | Proteine, die andere Proteine bei der Faltung unterstützen oder die Aggregat-bildung fehlgefalteter oder noch nicht komplett gefalteter Proteine verhindern. Die meisten werden als Hitzeschock-proteine (Hsp) bezeichnet, da sie bei erhöhter Temperatur vermehrt exprimiert werden. |
| Dies ist sinnvoll, da... Was passiert, wenn die Faltung misslingt? | ...Stressbedingungen die Effizienz der Proteinfaltung mindern. Misslingt eine korrekte Proteinfaltung, so werden die unbrauchbaren Proteine am Proteasom einem proteolytischen Abbau zugeführt. |
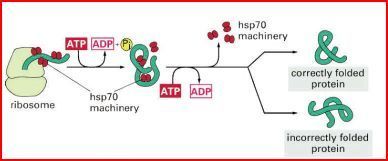
| Nach dem Austritt einer hydrophoben Aminosäure aus dem Ribosom... | ...wird diese von einem Chaperon abgedeckt. Nach der Fertigstellung des gesamten Proteins lösen sich alle hsp70 von den hydrophoben Aminosäuren, so dass sich das Protein im Idealfall richtig falten kann. |
| Funktion von Hsp70 | |
| Proteine mit einem komplizierten Faltungsweg müssen... | ...noch weiter "beschützt" werden: Sie gelangen in eine Proteintonne aus hsp60-Chaperonen. In diese Tonne passt nur ein ungefaltetes Protein, das so in "Isolationshaft" nicht mit anderen ungefalteten Proteinen aggregieren kann. |
| Die Tonne kann jetzt... | zusammen mit ihrem Inhalt ihre Form verändern, so dass die Proteine im Idealfall in ihre richtige Konformation "massiert" werden. Sowohl die hsp70- als auch die hsp60-Proteine verbrauchen viel Energie, die sie durch die Hydrolyse von ATP gewinnen. |
| Molekulare Chaperone des Hsp70-Typs unterstützen ... | die Proteinfaltung, indem sie diese Bereiche binden und eine Aggregation verhindern. Durch eine Folge von Zyklen der Bindung von Hsp70, Konformations- änderung mittels ATP-Hydrolyse und Freisetzung des Proteins wird ein korrekt gefaltetes Protein hervorgebracht. |
| Eine weitere wichtige Funktion von Hsp70 liegt... | ...im Transport von Proteinen in das Innere der Mitochondrien. Nur ungefaltete Proteine können durch Membranen gelangen. Durch Bindung an Hsp70 wird eine frühzeitige Faltung verhindert, bis das Protein transloziert werden kann. |
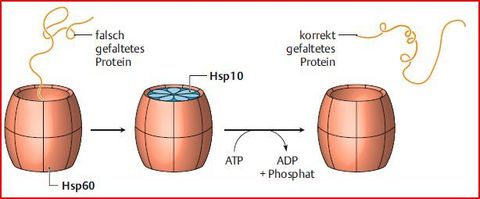
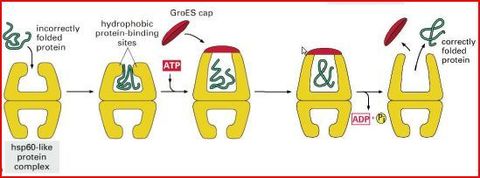
| Was können Hsp60 noch? | Fehlfaltung korrigieren. Wie bei Hsp70 erfolgt Erkennung/Bindung an Proteine über exponierte hydrophobe Oberflächen. Hat 14 identischen UE, die zwei heptamere, übereinander gestapelte Ringe bilden. Fassartige Struktur wird durch einen „Deckel“ aus weiteren heptameren Ring = Co-Chaperon Hsp10 abgeschlossen. |
| Funktion von Hsp60 bzw. Hsp10 | |
| Funktion von Hsp60 bzw. Hsp10 | |
| Was passiert im Inneren? | Fehlgefaltete Proteine gelangen in das Innere des „Fasses“ und erhalten dort unter ATP-Verbrauch eine andere Konformation |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Want to create your own Flashcards for free with GoConqr? Learn more.