4264159
Description
Flashcards by Mitra Shadloo, updated more than 1 year ago
|
|
Created by Mitra Shadloo
over 8 years ago
|
|
| Question | Answer |
| Die Reaktionsgeschwindigkeit | ist definiert als Änderung einer Konzentration pro Zeiteinheit. Ihre Einheit ist Mol pro Liter pro Sekunde. |
| Nimmt z. B. die Konzentration eines Stoffes A in einer Sekunde um 2 mmol/l ab, beträgt die Reaktionsgeschwindigkeit: | Das negative Vorzeichen zeigt das Absinken der Konzentration an. |
| Die Reaktionsgeschwindigkeit hängt ab von | 1. der Temperatur und 2. der Anfangskonzentration des Edukts bzw. der Edukte: Je höher seine/ihre Anfangskonzentration, desto größer ist die Reaktionsgeschwindigkeit. |
| Warum wird Pro Zeiteinheit wird nur ein bestimmter Prozentsatz der Ausgangssubstanz(en) zum Produkt umgesetzt? | Weil Moleküle nur unter bestimmten Bedingungen miteinander reagieren. So reagieren Moleküle in einer Lösung nur dann miteinander, wenn sie in einer bestimmten Orientierung und mit hinreichender Kraft zusammenstoßen. |
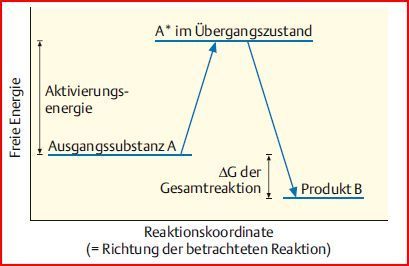
| Übergangszustand | Die Konfiguration, in der Moleküle miteinander reagieren, um eine neue chemische Verbindung zu bilden, bezeichnet man als Übergangszustand. Dieser Zustand ist energiereicher als der Ausgangszustand der Eduktmoleküle |
| Aktivierungsenergie | Die Energiedifferenz zwischen Ausgangs- & Übergangszustand der Edukte. Sie muss der Ausgangssubstanz zugeführt werden, damit diese in den Übergangs-zustand eintreten & Reaktion ablaufen kann. Ohne Aufwendung keine Reaktion, auch wenn das ΔG der Gesamtreaktion negativ ist. |
| Darstellung des Übergangszustandes im Energiediagramm | |
| Eine Reaktion läuft umso schneller ab, je... | ...mehr Moleküle einer Ausgangssubstanz pro Zeiteinheit den Übergangszustand durchlaufen, je niedriger also die Aktivierungsenergie ist. |
| Mitunter kann die Bildung eines Übergangszustandes durch die Wechselwirkung mit einem weiteren Stoff erleichtert werden. | Derartige Stoffe bewirken damit eine Beschleunigung der Reaktion. Sie werden allgemein als Katalysatoren bezeichnet. |
| Enzyme sind... | ...biologische Katalysatoren. Sie können Prozesse nur beschleunige. Sie können unmögliche nicht möglich machen. Allenfalls: energetische Kopplung herstellen, d. h. Prozess, der isoliert unmöglich wäre, mit anderem Prozess koppeln, um dann beide Prozesse gemeinsam ablaufen zu lassen. |
| Enzyme erleichtern | Molekülen den Eintritt in den Übergangszustand und senken auf diese Weise die Aktivierungsenergie. Dadurch erhöhen sie die Geschwindigkeit biochemischer Reaktionen. |
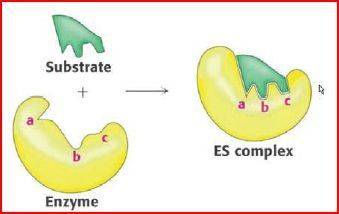
| katalytische (aktive) Zentren | Die Stellen, an denen Enzyme ihre Substrate binden und an denen die vom Enzym katalysierte Reaktion stattfindet, heißen katalytische (aktive) Zentren. zeigen eine hohe Substratspezifität. Sie bringen die Substrate in die optimale Reaktionsposition und erleichtern ihnen so den Eintritt in den Übergangszustand |
| Diese Bindung beruht auf dem... | "Schlüssel-Schloss-Prinzip", Substrat & Enzym haben im ES-Komplex zueinander passende Oberflächen. die Form kann sich aber während der Bindung sowohl des Enzymes als auch des Substrates verändern, was man als "induced fit" bezeichnet |
| "Induced fit" bei der Bildung des Enzym-Substrat-Komplexes | |
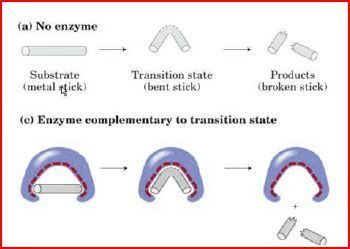
| Die Katalyse der chemischen Reaktion kommt dadurch zu Stande, dass... | ....das Enzym den Übergangszustand der Reaktion stabilisiert. Das bedeutet, dass nicht Substrat und Enzym, sondern Übergangszustand und Enzym besonders gut zueinander passen |
| der energiereiche Übergangszustand (gebogener Stab) wird durch die freiwerdende Bindungsenergie an das Enzym (hier magnetische Anziehung) begünstigt wird. | |
| Die Stabilisierung des Übergangszustandes gilt für... | ...beide Richtungen der Reaktion, die Hin- und die Rückreaktion werden prinzipiell in gleichem Maße beschleunigt: Kein Enzym kann aus Wasser und Kohlendioxid (ohne weitere Substrate) Glucose herstellen! Ein Enzym kann also nicht das Gleichgewicht einer Reaktion verschieben, sondern nur dessen Einstellung beschleunigen. |
| Enzyme sind... Affinität ist... | ...sowohl substratspezifisch als auch reaktionsspezifisch. Aufgrund der komplexen Oberfläche im aktiven Zentrum können die Enzyme meist sehr genau zwischen unterschiedlichen Substraten unterscheiden. ...die Stärke der Bindung eines Substrates an das Enzym. |
| Warum finden nur selten Nebenreaktionen statt? | Weil die Form des Übergangszustandes, der von einem Enzym stabilisiert wird, spezifisch für eine bestimmte Reaktion ist. |
| Was sind Coenzyme? | Im Gegensatz zu prosthetischen Gruppen sind Coenzyme funktionell Cosubstrate, da sie während der Reaktion umgewandelt werden. Sie dienen dem Austausch von Energie, Elektronen oder reaktiven Kohlenstoffverbindungen zwischen unterschiedlichen Enzymen. |
| Wo kommen sie vor? Was sind Beispiele für Coenzyme? | Jedes Coenzym kommt in vielen Stoffwechselreaktionen vor und ist damit so etwas wie eine "Währung" der Zelle. Beispiele sind die Folsäure, ATP und NADH. |
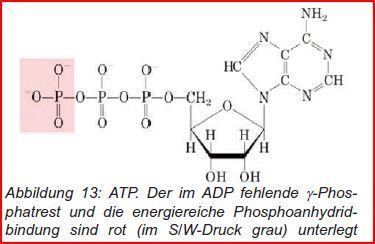
| Adenosintriphosphat (ATP) | Nukleotidtriphosphat mit der Stickstoffbase Adenin, das auch für die Biosynthese von Nucleinsäuren benutzt wird. In einigen Fällen werden auch UTP, GTP oder CTP als Energieüberträger im Stoffwechsel genutzt. |
| Im Körper sollen viele Reaktionen mit stark positivem dG ablaufen, bei denen im Gleichgewicht fast kein Produkt vorliegt: Beispiel: Phosphorylierung von Glucose Glucose+Pi <=> Glucose-6-Phosphat+H2O | Durch die Kopplung der endergonen Reaktion mit der stark exergonen Hydrolyse von ATP, bewerkstelligt im aktiven Zentrum des Enzyms Hexokinase, erreicht der Körper also die sehr effiziente Umwandlung der Glucose. |
| Nicotinamid-Adenin-Dinukleotid (NAD): Aus Nukleotiden & den Stickstoffbasen Adenin & Nicotinamid (=> aus Vitamin B3). Letzteres kommt in einer oxidierten (NAD+) & mit 2 e- extra in einer reduzierten Form (NADH) vor, die auch noch ein Proton aufgenommen hat . NADH hat ein sehr negatives Redoxpotential. | |
| Eigenschaften & Funktion von NADH/H+ | Ist bestrebt, die aufgenommenen e- wieder abzugeben, da diese sehr energiereich sind. Ähnliches gilt für NADP & FAD. Alle 3 werden daher im Stoffwechsel genutzt, um energiereiche e-, die bei Oxidationen entstehen, zu den Reduktionen, zu bringen => Daher Reduktionsäquivalente |
| Isoenzyme | Unter Isoenzyme versteht man unter- schiedliche Enzyme, die dieselbe Reaktion katalysieren aber verschiedene enzym-kinetischen Eigenschaften haben. Werden von unterschiedlichen Genen abgelesen & besitzen somit unterschiedliche, aber oft verwandte AS-Sequenzen. |
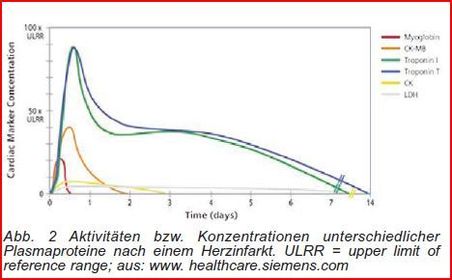
| Diagnostik mit Isoenzymen LDH | Beim Infarkt sterben Herzmuskelzellen, dadurch gelangen intrazelluläre Enzyme ins Blut. Auftreten & Menge der Enzyme im Blut deuten auf die Schwere hin. Die LDH & CK kommen auch außerhalb des Herzens vor. Unterscheidung, durch andere Isoformen dieser Enzyme im Herzen. |
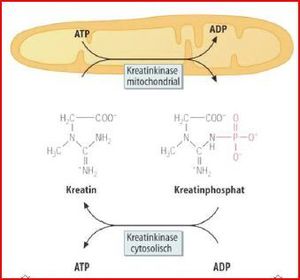
| Kreatinkinase | ATP im Muskel ist bei Belastung nach ~2-3 s verbraucht. Um zusätzlich Energie zu speichern, gibt es im Muskel Kreatin-phosphat, kann aus Nahrung kommen/mit geringerem Aufwand als ATP vom Körper hergestellt werden. CK macht in Ruhe aus ATP & Kreatin Kreatinphoshat |
| Kreatinkinase | |
| CK-Isoformen | CK hat eine Quartärstruktur & kommt in mehreren Isoformen vor. cytosolische Enzym = Dimer, aus B- und M-UE besteht. CK-1 (CK-BB) => überwiegend im Gehirn, CK-3 (CK-MM) hauptsächlich im Herz- und Skelettmuskel. Konzentration von CK-2 (CK-MB-Heterodimer) ist hingegen im Myokard am höchsten. |
| Die CK-Menge kann am einfachsten durch Messung der Aktivität im Enzymtest bestimmt werden. Die Gesamt-CK-Aktivität ist normalerweise auf die CK-MM Isoform zurückzuführen. Bei einem Infarkt wird aus den geschädigten Herzmuskelzellen zusätzlich CK-MB freigesetzt | |
| Regulierbarkeit der Enzyme | Je nachdem, ob in einer Stoffwechselsituation eine bestimmte Reaktion benötigt wird oder nicht, kann das zugehörige Enzym aktiv oder inaktiv sein. |
| Schrittmacherenzyme & Endprodukthemmung | Die Schrittmacherenzyme, die die erste spezifische Reaktion für einen Synthese-weg katalysieren, werden häufig durch die Endprodukte der Synthesewege gehemmt (Endprodukthemmung). So können die Konzentrationen von Metaboliten relativ konstant gehalten werden. |
| Endprodukthemmung: | Hemmung eines (Schrittmacher- oder Schlüssel-) Enzyms durch das Endprodukt zur negativen Rückkopplung |
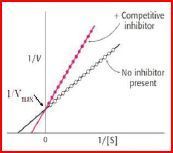
| Kompetitive Hemmung: | häufigsten Form der reversiblen Enzym-hemmung, der Inhibitor konkurriert mit dem natürlichen Substrat um die Bindung an das Enzym. Dadurch wird die katalytische Aktivität des Enzyms herab-gesetzt. Sie kann jedoch durch Zugabe größerer Mengen des natürlichen Substrats wieder gesteigert werden. |
| Was passiert mit vmax? Was passiert mit Km? | In Gegenwart von hinreichend großen Konzen. des Substrats kann sogar der ursprüngliche vmax-Wert erreicht werden. vmax also = unverändert! Der Km-Wert wird durch Zugabe des Inhibitors hingegen erhöht. |
| Warum wird Km erhöht? | Der Km-Wert gibt an, welche [Substrat] erforderlich ist, damit die Hälfte der Substratbindestellen der Enzymmoleküle mit Substrat besetzt sind. In Gegenwart eines kompetitiven Inhibitors ist aber eine erhöhte [Substrat] erforderlich, um die Hälfte der Bindestellen der Enzymmoleküle besetzen zu können. |
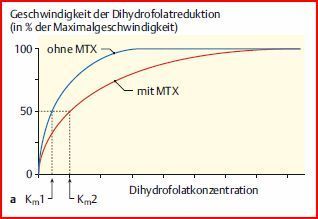
| Methotrexat | u. a. in Krebstherapie eingesetzt. hemmt Dihydrofolat-Reduktase kompetitiv. Reduziert Dihydrofolat zu Tetrahydrofolat, das eine wichtige Rolle im Stoffwechsel der Nukleotide – und damit bei der DNA-Synthese – spielt. Die Tumorzellen bilden daraufhin weniger Tetrahydrofolat und ihr Wachstum wird gehemmt. |
| Effekt des kompetitiven Inhibitors Methotrexat (MTX) auf die Dihydrofolat- Reduktase (DHFR) 1) Darstellung im Michaelis- Menten-Diagramm. | |
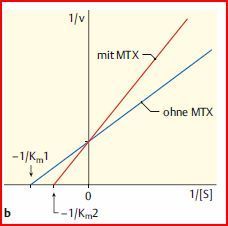
| Darstellung im Lineweaver- Burke-Diagramm. | |
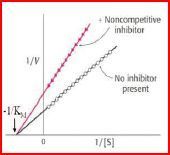
| Nicht-kompetitive Allosterische Hemmung | Inhibitoren, die Aktivität des Enzyms unterdrücken,indem sie an eine Bindungs-stelle außerhalb der Substratbindungs-stelle binden. Die Hemmung kann dann durch höhere Substratkonzentrationen nicht mehr rückgängig gemacht werden, vmax ist erniedrigt, die Hemmung ist nicht kompetitiv. Km unverändert. |
| Beispiel nicht-kompetitive Hemmung | Schwermetalle entfalten ihre giftige Wirkung über ihre Bindung an SH-Gruppen von Enzymen. |
| Funktionsweise von Medikamenten als Enzyminhibitoren Bsp.: Penicillin | Hemmt Zellwandsynthese der Bakterien, ähnelt dem D-Ala-D-Ala-Peptid, das von der Glykopeptidtranspeptidase zum Quervernetzen des Mureinsacculus benutzt wird. Allerdings besitzt es den ß- Laktam-Ring = zweite Bindung zwischen den imitierten AS. |
| Was passiert jetzt? | Anlagerung in das aktive Zentrum wird wie bei dem Peptid eine Bindung zwischen den "AS" gespalten. Da der zweite Teil des ß-Laktam-Ringes noch intakt ist, bleibt das Penicillin irreversibel kovalent am Enzym gebunden. Wie beim Aspirin kommt es hier zur Enzymvergiftung durch einen Selbstmordinhibitor, |
| Arsen | Arsen in Form von Arsenit reagiert z.B. mit den SH-Gruppen der Dihydrolipon-säure. Diese prosthetische Gruppe des Pyruvatdehydrogenasekomplexes kann da durch nicht mehr regeneriert werden. Der Komplex ist vor allem für Nervenzellen wichtig, die als Hauptenergiequelle Glukose benutzen. |
| Cyanid Was macht es? | Cyanid (Blausäure), das an Fe3+ in Hämkomplexen bindet = prosthetische Gruppen von O2-bindenden Proteinen, zum einen in der Cytochromoxidase (katalysiert im letzten Schritt der Atmungskette die Entstehung von Wasser aus Sauerstoff) und zum anderen im Hämoglobin. |
| Was ist die Folge? | Die Inhibition der Cytochromoxidase führt dazu, dass in den Mitochondrien kein ATP mehr synthetisiert werden kann und die Zelle stirbt. Als Therapie bei einer Cyanidvergiftung kann man Stoffe wie Nitrit spritzen, die das Hämoglobin (enthält Fe2+ im Häm) im Blut zum Teil in Methämoglobin (mit Fe3+) umwandeln. |
| Warum Methämoglobin? | Das Methämoglobin kann zwar keinen Sauerstoff mehr transportieren, bindet aber dafür das Cyanid mit höherer Affinität als die Cytochromoxidase. Nicht bei CO-Vergiftung geben! Da sind viele Hämoglobine schon durch CO blockiert! |
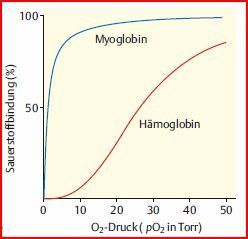
| Allosterische Effekte | beruhen auf strukturellenVeränderungen eines Proteinmoleküls, die sich innerhalb des Moleküls über eine größere Distanz fortpflanzen. Geschieht das über mehrere UE = Kooperativität Bsp.: Hämoglobin. wichtige Voraussetzung für die Möglichkeit, die Eigenschaften von Proteinen/Enzyme zu modulieren. |
| Kooperativität | Eine weitere Möglichkeit der Regulation für Proteine mit einer Quartärstruktur,: Kommunikation zwischen den UE = kooperativität Bsp: Hämoglobin. Hier findet "positive Kooperativität" statt & bewirkt, dass O2 sehr viel effizienter von der Lunge zu den anderen Organen transportiert werden kann. |
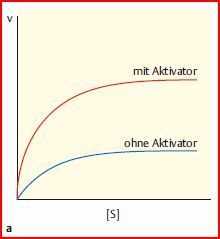
| Allosterische Effekte | Außerhalb des katalytischen Zentrums enthalten viele Enzyme eine Bindestelle für einen Metaboliten, der die Aktivität des Enzyms als Aktivator stimuliert bzw. als Inhibitor hemmt. Die Bindung eines Aktivators kann z. B. eine Erhöhung der maximalen Reaktionsgeschwindigkeit vermitteln |
| Steigerung der maximalen Reaktionsgeschwindigkeit durch einen allosterischen Aktivator (allosterische Regulation vom V-Typ). Erhöhung von vmax | |
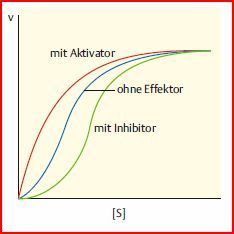
| Beeinflussung der Enzymaffinität durch allosterische Effektoren (allosterische Regulation vom K-Typ). Änderung Km-Wert | |
| Dagegen Myoglobin | Das O2- bindende Protein Myoglobin hingegen, das in Skelett- und Herzmuskelzellen als O2-Reservespeicher bei ungenügendem O2-Angebot dient, liegt nur monomer (1 UE) vor. Deshalb zeigt es bei steigenden [O2] keine Kooperativität, sondern lediglich eine einfache Zunahme der Sauerstoffbindung. |
| O2-Bindungskurven von Hämoglobin und Myoglobin: Anders als bei Myoglobin wirkt O2 bei Hämoglobin als allosterischer Aktivator. Die Bindung von O2 an eine Hämoglobin-Untereinheit steigert die O2-Affinität der übrigen Untereinheiten | |
| Regulation durch Phosphorylierung | Um aber auf veränderte Umwelt-bedingungen zu reagieren, muss der Körper auch unabhängig von der [Produkt] Stoffwechselwege regulieren können. Diese übergeordnete Regulation wird meist von Hormonen gesteuert, die als Signale viele Zellen beeinflussen. |
| Wie und wo geschiet das? | Durch unterschiedliche Signalkaskaden, werden die Enzyme in der Zelle gesteuert. Auf der molekularen Ebene geschieht das häufig durch eine Phosphorylierung. Hierbei werden Phosphatreste von ATP (siehe unten) auf OH-Gruppen von Tyrosin-, Serin- oder Threoninresten übertragen. |
| Was passiert dadurch? | Ähnlich wie bei den allosterischen Inhibitoren bewirkt die Anheftung des negativ geladenen Phosphates eine Konformationsänderung des aktiven Zentrums, die jetzt zu einer Inhibition oder aber auch zu einer Aktivierung des Enzymes führt. |
| Zymogenaktivierung | Proteasen aus Pankreas wie Trypsin & Chymotrypsin, die Nahrungsproteine im Darm verdauen sollen, dürfen nicht die Pankreasproteine selbst angreifen. Darum werden sie als inaktive Vorstufen (Zymogene) synthetisiert und erst im Darm selbst aktiviert. |
| Wie wird aktiviert? | Dies geschieht durch Proteolyse der Protease selbst: Ein anderes, schon aktives Proteasemolekül schneidet eine ganz bestimmte Peptidbindung des Zymogens. Dadurch kommt es wiederum zu einer Konformationsänderung und damit Aktivierung des aktiven Zentrums. |
| Wo findet was ähnliches statt? | Eine ähnliche Prozessierung kennt man beim Kollagen des Bindegewebes und vielen Faktoren des Blutgerinnungssystems. |
| Michaelis-Menten-Kinetik | Im einfachsten Fall einer Michaelis-Menten-Kinetik katalysiert ein Enzym E die Umwandlung eines Substrats S in das Produkt P. Dabei bildet sich zunächst ein Enzym-Substrat-Komplex ES. |
| Was passiert dann? | Substrat durchläuft am Enzym den Übergangszustand, reagiert zu Produkt P &verlässt Enzym. Im einfachsten Fall – wenn das Produkt P zum Enzym nur eine geringe Affinität hat oder wenn die Konzentration des Produkts sehr gering ist – kann die Rückreaktion von P zu S zunächst vernachlässigt werden. |
| Reaktionsschema | |
| Der geschwindigkeitsbestimmende Schritt unter diesen Voraussetzungen ist... | ...die Reaktion ES → E + P. Die Geschwindigkeit, mit der sich P bildet, ist also nur von der Geschwindigkeits-konstante k2 + [ES] abhängig. Die Geschwindigkeitskonstante k2 gibt dabei den Prozentsatz an ES an, der innerhalb einer Sekunde in E und P zerfällt. |
| Daraus ergibt sich eine Formel für die Reaktionsgeschwindigkeit: | |
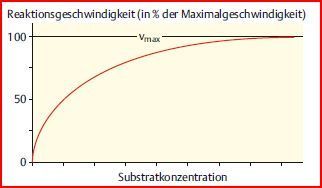
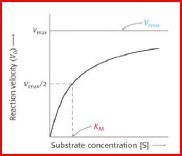
| Wie lässt sich die Reaktionsgeschwindigkeit steigern? | Die Reaktionsgeschwindigkeit lässt sich durch Erhöhung der [Substrat] steigern, nur bis zu einem Maximalwert = bis zu der Konzentration, wo alle Enzyme = ES, bei der das Enzym also gesättigt ist. Bei dieser [Substrat] läuft die Reaktion mit der max. Geschwindigkeit vmax ab. |
| Stellt man die Reaktionsgeschwindigkeit in Abhängigkeit von der [Substrat] grafisch dar (Michaelis-Menten-Diagramm), ergibt sich eine Hyperbel, die der maximalen Geschwindigkeit vmax entgegenstrebt | |
| Fazit | Die Maximalgeschwindigkeit vmax einer enzymkatalysierten Reaktion hängt von der Enzymkonzentration ab. |
| Manche Medikamente bewirken eine irreversible Inaktivierung bestimmter Enzyme. Beispiel ASS | Acetylsalicylsäure inaktiviert das Enzym Zyklooxygenase, das bei der Synthese von Prostaglandinen aus Arachidonsäure eine zentrale Rolle spielt. Dadurch sinkt die Konzentration der funktionsfähigen Enzyme & damit die max. erreichbare Konzentration des ES. & dann auch vmax. |
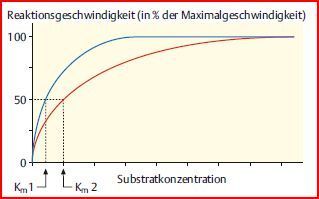
| Die Michaelis-Menten-Konstante Km | Km ist diejenige [Substrat], bei der die Hälfte der Enzyme mit Substrat beladen ist. Bei dieser [Substrat] beträgt die Reaktions-geschwindigkeit vmax/2. Da es sich bei der Michaelis-Menten-Konstante um eine bestimmte [Substrat] handelt, hat sie die Einheit Mol/Liter. |
| Michaelis-Menten-Diagramm zweier Enzyme gleicher Substratspezifität, aber mit unterschiedlichem Km-Wert | |
| Km1 | Das Enzym mit dem niedrigeren Km-Wert ist bereits bei niedrigen [Substrat] zur Hälfte mit Substraten beladen. Es zeigt also eine große Bereitschaft, seine Substrate zu binden: Das Enzym zeigt zu seinen Substraten eine hohe Affinität |
| Km2 | Das Enzym mit dem höheren Km-Wert ist erst bei deutlich höheren [Substrat] zur Hälfte beladen, hat also eine geringere Affinität zu seinen Substraten. Dieses Enzym setzt also bei niedrigen [Substrat] weniger Substrat um als das Enzym mit hoher Affinität. |
| Fazit | Die Michaelis-Menten-Konstante (der Km-Wert) ist ein Maß für die Affinität eines Enzyms zu seinen Substraten: Je kleiner Km, desto höher die Affinität. |
| Wovon hängt die Affinität eines Enzyms zu seinen Substraten ab? | Sind die Reaktionspartner im Reaktions-schema im chemischen Gleichgewicht, ist die Affinität umso höher, je mehr ES vorliegen. Die Zahl der ES ist umso größer, je schneller sich ES aus E und S bildet und je langsamer ES in E + S bzw. in E + P zerfällt. |
| Es lässt sich zeigen, dass der Km-Wert... | ...unmittelbar aus dem Verhältnis der zugehörigen Geschwindigkeitskonstante berechnet werden kann. |
| Wie ändert sich der Km-Wert, wenn die Enzymkonzentration und mit ihr der vmax- Wert reduziert wird, z. B. bei der Zyklooxygenase durch die Reaktion mit Acetylsalicylsäure? | Da die relevanten Geschwindigkeitskonstanten k-1, k2 und k1 dabei unverändert bleiben, ändert sich die Affinität der übrig gebliebenen Enzyme zu ihren Substraten nicht. Der Km-Wert bleibt trotz Reduktion des vmax-Wertes unverändert. |
| Fazit | Die Michaelis-Menten-Konstante ist von der Enzymmenge unabhängig |
| Die Michaelis-Menten-Gleichung | Sind vmax und der Km-Wert eines Enzyms bekannt, kann man für jede [Substrat] [S] die entsprechende Reaktionsgeschwindigkeit v berechnen. Hierzu setzt man diese Werte in die Michaelis-Menten-Gleichung ein, die die Hyperbel des Michaelis-Menten-Diagramms beschreibt |
| Stellt man sie etwas anders dar, kann man sich einige Charakteristika enzymkatalysierter Reaktionen ableiten: | Bei [S] >> Km ist v = vmax. Bei [S] = Km ist v = vmax/2. Bei geringen [Substrat] (also einer hohen Konz. an freiem Enzym): Verdopplung der [Substrat]=>Verdopplung der RG. Bei höheren [Substrat] gibt es jedoch immer weniger freie Enzyme, so dass schließlich alle Enzyme mit Substrat gesättigt sind und v Vmax erreicht. |
| Substratsättigungsdiagramm | |
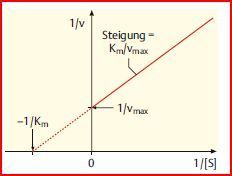
| Lineweaver-Burk- Diagramm | Darüber hinaus muss vmax bekannt sein, um die Michaelis-Menten-Konstante Km bestimmen zu können, denn diese Konstante ist über die halb-vmax definiert. Leider ist es sehr schwierig, vmax aus einem Michaelis-Menten-Diagramm abzulesen, daher trägt man alternativ 1/V gegen 1/[S] auf |
| Lineweaver-Burk- Diagramm | In diesem Diagramm ist die obige Kurve in eine Gerade umgewandelt. Der Abschnitt auf der Y-Achse entspricht 1/Vmax, der Schnittpunkt mit der X-Achse -1/KM, so dass diese Werte leicht abgelesen werden können. |
| Lineweaver-Burk-Diagramm | |
| Methanolvergiftung | Die Alkoholdehydrogenase: macht aus Methanol Formaldehyd, das schnell zu Ameisensäure weiterreagiert, die die Cytochromoxidase in den Mitos hemmt. Da vor allem die Retina sehr empfindlich reagiert, kann es zur Erblindung kommen. Ethanol hat höhere Affinität. Ist jedoch zuviel Methanol vorhanden: Vergiftung |
| Therapie | Zur Therapie stehen dem Arzt zwei Inhibitoren der Alkoholdehydrogenase zur Verfügung: Klassischer weise werden hohe Dosen Ethanol verabreicht, was zu einer Verlangsamung des Methanolabbaus und damit der Ausscheidung |
| Koffein | Coffein = Purin ähnlich dem im Nukleosid Adenosin vorkommenden Adenin. Adenosin bindet als Hormon im Gehirn löst inhibitorische Signale aus=> Müdigkeit & Schlaf+verminderter Ausschüttung Adrenalin/Dopamin, vermindert Wirkung von Adenosin, indem es als Antagonist an die Rezeptoren bindet. |
| Wirkung | Wachheit und erhöhte Ausschüttung von Adrenalin (führt zu schnellerem Herzschlag und anderen stoffwechselaktivierenden Effekten) und von Dopamin (Neurotrans- mitter, der ein Glücksgefühl auslöst) |
| Zweite Wirkung | Ein zweiter Angriffspunkt für Coffein ist eine Isoform der Phosphodiesterase. Dieses Enzym baut unter anderem cAMP (cyclisches Adenosinmonophosphat), das das Adrenalinsignal in der Zelle weiterleitet, ab. In hohen Konz. bewirkt Koffein: doppelt gesteigerten Adrenalinwirkung im Körper |
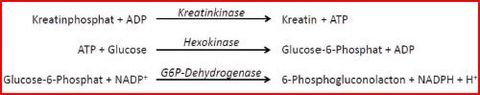
| Die Bestimmung der CK-Aktivität erfolgt durch ... | ...einen gekoppelten enzymatischen Test. Das in der CK-Reaktion entstehende ATP wird in einer stark exergonen Reaktion praktisch vollständig genutzt, um Glucose zu phosphorylieren. In 3. Reaktion reagiert dann Glucose-6-Phosphat unter Bildung von einem NADPH-Molekül pro umgesetztem CK-Molekül |
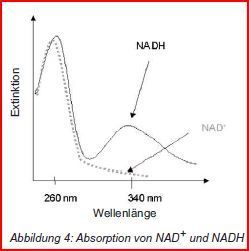
| Warum nutzen wir drei Reaktionen? | Die Messung der Produktentstehung erfolgt am einfachsten im Photometer. Allerdings absorbieren Kreatin und ATP in sehr ähnlicher Weise Licht wie CK & ADP. Darum: In Enzymtests werden weitere Reaktionen an den 1. Reaktionsschritt "gekoppelt", die letztendlich: Umwandlung von NAD(P)+ in NAD(P)H führen. |
| Warum NAD(P)+ / NAD(P)H/H+? | Diese absorbieren UV-Licht einer Wellenlänge von 340 nm unterschiedlich stark |
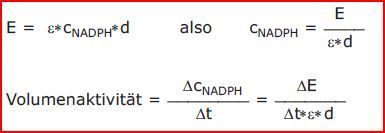
| Berechnung der Volumenaktivität | Man untersucht wie viel Substrat von einer Serumprobe pro Zeit unter optimalen Bedingungen bei Substratsättigung umgesetzt wird. Nehmen Sie hierzu den ermittelten Vmax-Wert, dieser entspricht DE/Dt. Die Volumenaktivität ist die Enzymaktivität pro Volumen. |
| katalytische Aktivität | Die katalytische Aktivität eines Enzyms lässt sich nur berechnen, wenn sowohl der vmax als auch der Km-Wert bekannt sind. 1 Unit (U) ist die katalytische Aktivität derjenigen Enzymmenge, die bei 30°C 1 μmol Substrat pro Minute unter optimalen pH-Bedingungen und bei Sättigung mit Substrat umsetzt. |
| Um die Volumenaktivität aus der Reaktionsgeschwindigkeit zu berechnen, müssen Sie mit Hilfe des Lambert-Beerschen-Gesetzes die Konzentration von NADPH aus dessen Extinktion berechnen: | |
| Wechselzahl | Unter der Wechselzahl versteht man die Anzahl der pro Mol Enzym in einer Zeiteinheit umgesetzten Mole Substrat. Diese Zahl ist identisch mit der pro Enzymmolekül in einer Zeiteinheit umgesetzten Substratmoleküle. |
| Um die Wechselzahl zu bestimmen, ermittelt man... | ...vmax für eine definierte Menge an Enzym. |
| Grundsätzlich sind zwei Wege zu unterscheiden, über die ein Signal in die Zelle gelangen kann: 1) | I) das Hormon wird durch die Plasma membran in das Innere der Zelle transportiert, wo es entweder im Cytosol oder im Zellkern als Ligand an einen spezifischen Hormonrezeptor bindet. |
| 2) | II) nicht das Hormon gelangt in die Zelle, sondern nur das Signal. In diesem Fall bindet das Hormon an die extrazelluläre Domäne eines in der Plasmamembran verankerten spezifischen Hormonre - zep tors, der das Signal durch kon for - mationsänderung des Transmembran - pro teins ins Innere der Zelle weiterleitet. |
| Zur zweiten gruppe gehören alle... | hydrophilen Signalmoleküle ein schließ lich der neurotransmitter, Protein hormone und Wachstumsfaktoren sowie einige hydrophobe Hormone wie die eikosanoide (Prostaglandine, Thromboxane usw). Sie alle binden im Allgemeinen mit hoher Affinität als Liganden an spezifische Zelloberflächenrezeptoren. |
| Je nach dem Mechanismus, über den die Weitergabe des Signals erfolgt, unterscheidet man 4 verschiedene Rezeptortypen: | 1. Ionenkanäle 2. G-Protein gekoppelte Rezeptoren 3. Rezeptoren mit eigener Enzymaktivität 4. Rezeptoren, die mit Enzymen assoziiert sind. |
| G-Protein gekoppelte Rezeptoren | gehören zu einer Familie homologer Proteine, die die Plasmamembran 7x durchspannen. Wirken indirekt, indem sie die Aktivität eines weiteren membran-gebundenen Zielproteins, das ein Enzym oder Ionenkanal sein kann, regulieren. |
| Wie wird die Signalübertragung vermittelt? | Die Signalübertragung zwischen Rezeptor und Zielprotein wird durch ein drittes Protein, ein heterotrimeres GTPbindendes Regulatorprotein (G-Protein) vermittelt. Wenn das Zielprotein ein Enzym ist, wird die Konzentration intrazellulärer Media-toren - genannt "second messenger" - verändert. |
| Die Konzentration folgender intrazellulärer Mediatoren wird über G-Proteingekoppelte Rezeptoren beeinflusst: | cAMP-Zunahme durch Aktivierung der Adenylatcyclase, IP3 & Diacylglycerin(DAG)- Bildung durch Aktivierung von Phospholipase C, cGMP-Spaltung durch Phosphodiesterasen |
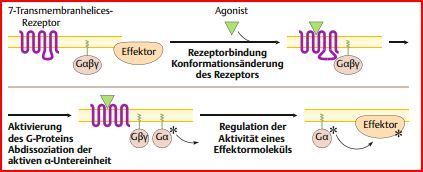
| Mechanismus der Signaltransduktion | Die Signalkette G-Protein-gekoppelter Rezeptoren ist aus drei Modulen aufgebaut: Rezeptor, heterotrimeres G-Protein (es gibt auch kleine G-Proteine), Effektormolekül (Enzym oder Ionenkanal) Nach Bindung des Liganden aktiviert der Rezeptor das G-Protein, welches wiederum ein Effektormolekül aktiviert. |
| Aufbau des Rezeptors Aufbau des Rezeptors Alle G-Protein-gekoppel | N-Terminus des Proteins liegt EX, der C-Terminus IZ Die Helices sind durch Schleifen verbunden. Liganden binden an EZ-Bereiche. Durch Bindung der Liganden kommt es wohl zu einer Konformations-änderung der Helices=>überträgt sich auf die Schleifen der IZ-Seite=>Bindungsstelle für G-Protein wird geschaffen |
| Schematische Darstellung der G-Protein-vermittelten Signaltransduktion | |
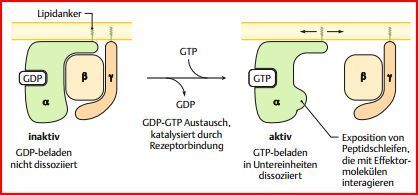
| Aufbau der heterotrimeren G-Proteine | aus drei mit α, β & γ bezeichneten UE aufgebaut. Die α-UE (Gα) hat ein GDP oder GTP gebunden. Die β- & die γ-UE bilden einen stabilen Komplex & sind über einen Lipidanker der γ-Untereinheit in der Membran verankert. Die α-UE auch & kann regulierbar mit der β/γ-UE und dem 7-TM-Rezeptor interagieren. |
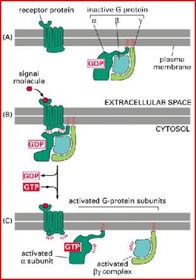
| In der nicht stimulierten Zelle: | In der nicht stimulierten Zelle bilden alle drei Untereinheiten einen inaktiven Komplex, der aber nur stabil ist, solange die α-Untereinheit GDP gebunden hat |
| Reaktionszyklus der heterotrimeren G-Proteine Aktivierung des G-Proteins: | Bindung eines EZ Signalmoleküls an den Rezeptor => Konformationsänderung (= Aktivierung) des Rezeptors=> Rezeptor kann die α-UE binden=> Konformations-änderung der α-UE => Abdissoziation von GDP => freie Bindungsstelle von GTP-eingenommen=> bindet & stabilisiert neue Konformation => G-Protein aktivieret |
| Die Konformationsänderung der α-Untereinheit nach Bindung durch den Rezeptor hat zwei wichtige Konsequenzen | 1. Die α-UE löst sich vom Rezeptor & der β/γ-UE, kann sich frei in Membran bewegen, durch Lipidankers nicht abdissoziieren. 2. In der neuen Konformation der α-UE sind Peptidschleifen exponiert, die mit dem Effektormolekül interagieren und dieses so aktivieren können |
| Inaktivierung des G-Proteins: | erfolgt durch die intrinsische GTPase-Aktivität der aktivierten Gα-UE: Gα hydrolysiert GTP langsam zu GDP und Phosphat. In der GDP-gebundenen Form assoziiert die α-UE wieder mit der β/γ-UE und ist dann bereit für eine erneute Aktivierung. |
| GTPase-aktivierender Proteine (GAPs) | Die Hydrolyse von GTP kann durch Bindung GAPs beschleunigt werden. Die bekanntesten sind die RGS(Regulator of G-Protein-Signaling)-Proteine, die die GTPase-Aktivität bis zu 1000-fach erhöhen können=wichtige Regulatoren der Aktivität von G-Proteinen, könnten Teil eines negativen Feedbacks sein |
| Die heterotrimeren G-Proteine werden nach der Familienzugehörigkeit der α-UE in vier Hauptklassen unterteilt: Gs | Gs: ubiquitär, aktivierende Liganden: Adrenalin (β-adrenerge Rezeptoren), Glucagon, TSH, LH, FSH, ADH. Effektorproteine: Adenylatzyklase ↑ |
| Gi | ubiquitär, aktivierende Liganden: Adrenalin (α2-adrenerge RZ), Effektorproteine: αi-UE: Adenylatzyklase ↓ β/γ-UE: Phospholipase Cβ ↑ – K+-Kanäle ↑ – Ca2+-Kanäle ↓ |
| Gq | ubiquitär, aktivierende Liganden: Adrenalin (α1-adrenerge Rezeptoren), Effektorproteine: Phospholipase Cβ ↑, IP3 ↑, DAG ↑, Ca2+ ↑ |
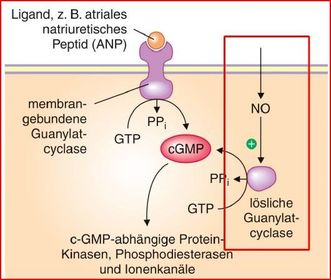
| Enzymgekoppelte Rezeptoren Guanylatzyklasen | Guanylatzyklasen sind Hormonrezeptoren, die bei Aktivierung den Second Messenger cGMP synthetisieren. |
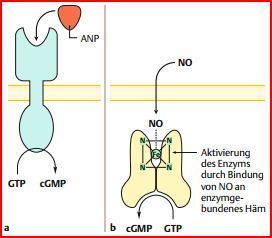
| Es gibt zwei Klassen von Guanylatzyklasen, die sich in ihrer Struktur und ihrer subzellulären Lokalisation, vor allem aber in ihrer Aktivierung durch Liganden unterscheiden: | 1) membrangebundene Guanylatzyklasen werden von extrazellulären Liganden aktiviert. Zu dieser Gruppe gehören z. B. Rezeptor für ANP & Guanylatzyklase, die das für den Sehvorgang wichtige cGMP bildet 2) Zytosolische (lösliche) Guanylatzyklasen werden durch NO aktiviert. |
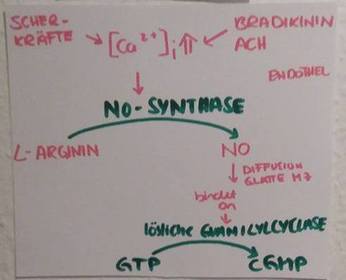
| cGMP-vermittelte Wirkungen Hauptfunktion von NO | NO diffundiert von seinem Bildungsort (z. B. Endothel- oder Nervenzelle) in die Zielzelle (z. B. glatte Muskelzelle, Thrombozyt, Nervenzelle) und aktiviert dort die zytosolische Guanylatzyklase, führt also zur Bildung von cGMP. Dies ist die Hauptfunktion von NO. |
| Die Folgen sind: | Relaxation der glatten Gefäßmuskulatur, Hemmung der Thrombozytenaggregation, Förderung der Signalübertragung an Synapsen des Gehirns. Diese ist für Lernprozesse wichtig. |
| NO führt über die Bildung von cGMP zur Relaxation der glatten Muskulatur | > Blutdrucksenkung durch Vasodilatation |
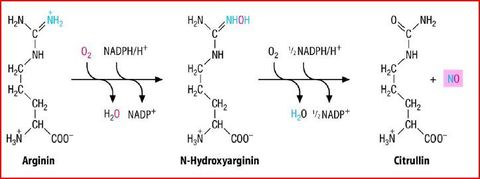
| NO - Synthase | |
| Guanylatcyclase: | lösliche, dimere Form, die durch NO aktiviert wird & membrangebundene Form, die durch ez Liganden aktiviert wird (z.B. ANP). katalysiert die Bildung von 3´, 5´-cyclo-GMP aus GTP unter Abspaltung von PP (analog der cAMP-Bildung), Abbau durch cGMP-Phosphodiesterase (cGMP-PDE) zu 5´-GMP |
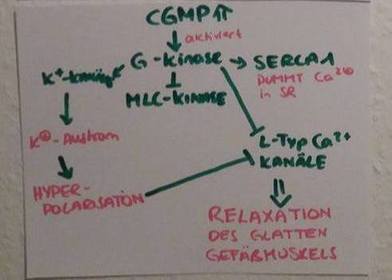
| cGMP aktiviert die Proteinkinase G (PKG) | - hoch exprimiert in glatter Muskulatur, Thrombocyten und Kleinhirn - relaxierende Wirkung durch Phosphorylierung und Aktivierung von Ca2+-ATPasen der Plasmamembran => Ca2+-Konzentration im Cytosol sinkt ) => Relaxation der Glattmuskelzelle |
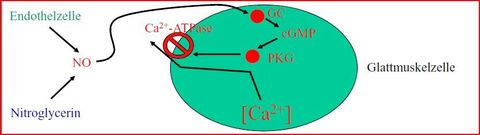
| Signalweg NO & Gefäße Teil 1 | Stimulation der Endothelzellen mit Acetylcholin oder Bradikinin zur Bildung von NO aus Arginin durch die NO-Synthase. Das NO diffundiert in benachbarte Endothelzellen & bindet dort an die lösliche Guanilatzyklase. Diese macht aus GTP=>cGMP |
| Signalweg NO & Gefäße Teil 2 | cGMP aktiviert die PKG=> diese hemmt die MLC-Kinase & die L-Typ Ca2+-Kanäle + Aktiviert K+-Kanäle, die K+ ausströmen lassen: es kommt zur Hyperpolarisation, was wiederum die L-Typ Ca2+-Kanäle hemmt + aktiviert die SERCA 1, die Ca2+ ins SR pumpt => zusammen [Ca2+]i sinkt => glatte Muskelzelle relaxiert. |
| Wie wird cGMP inaktiviert? | Ebenso wichtig wie die Bildung der beiden Second Messenger cGMP und cAMP ist deren Inaktivierung: Phosphodiesterasen bauen cGMP zu GMP und cAMP zu AMP ab |
| Für welchen Vorgang ist NO noch wichtig? Blutdruckregulation | Bradykinin bindet an Bradykinin-RZ im Gefäßendothel => über NO-Kaskase=> erhöhte Gefäßpermeabilität + Tonusänderung der glatten Muskulatur in der Gefäßwand. Im Effekt kommt es dadurch zu einer Vasodilatation und zu einem erhöhten Flüssigkeitsausstrom aus dem Gefäßsystem. |
| Für welchen Vorgang ist NO noch wichtig? Erektion | Aktivierung der Endothelzellen in den Blutgefäßen Klitoris/Penis ist wichtig => Aktivierung der endothelialen NO-Synthase => Botenstoff (NO), im entsprechenden Gefäßabschnitt freigesetzt => Aktivierung der Guanylylzyklase=>Bildung von cGMP) |
| Folge: | Durch die dann erfolgende Blutgefäßerweiterung füllen sich die klitoralen oder penilen Schwellkörper mit Blut (Vasokongestion) |
| Wie wirkt Viagra? | Sildenafil = PDE-Hemmer Typ V (cGMP-spezifisch). Durch die Hemmung=>cGMP langsamer abgebaut => Erektion dauert länger + generelle Gefäßdilatation => Blutdruck fällt um 10 mmHg (Gesunden). Gleichzeitige Einnahme von anderen gefäßerweiternden Medikamenten kann lebensgefährlich werden! |
| Sehvorgang | cGMP öffnet Kationenkanäle => Licht wird von Rhodopsin absorbiert => das über G-Protein Transducin die Phosphodiesterase aktiviert => Diese spaltet das cGMP, die Kanäle schließen, und der Photorezeptor hyperpolarisiert => Signal zum Gehirn: "Licht" |
| Nebenwirkungen Viagra | Hemmt auch die PDE im Auge (Hemmung nicht spezifisch genug!) => cGMP nicht genug abgebaut => Lichtsignale können nicht richtig verarbeitet & dann auch nicht mehr weitergegeben werden => Sehstörungen |
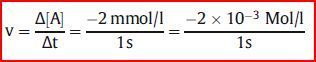{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
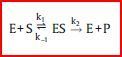{kind=link}
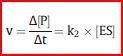{kind=link}
{kind=link}
{kind=link}
{kind=link}
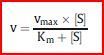{kind=link}
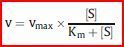{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Want to create your own Flashcards for free with GoConqr? Learn more.