37017534
Description
Mind Map by Vicky Tituaña, updated more than 1 year ago
|
|
Created by Vicky Tituaña
over 3 years ago
|
|
Metabolismo de Lípidos
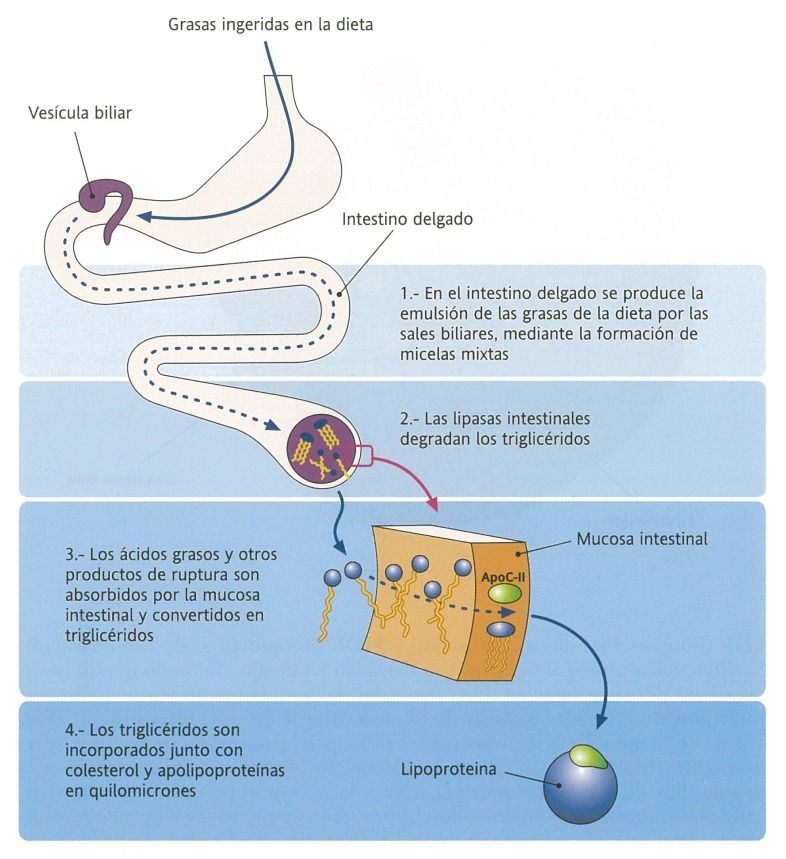
- Digestión y absorción
de lípidos
- Emulsión de las
grasas
ingeridas
- Los lípidos no pueden atravesar libremente las membranas de
las células epiteliales del intestino ni enterocitos, debido a la
presencia de una capa de agua que rodea las microvellosidades.
- Previamente a la digestión enzimática, debe producirse la emulsión
de las grasas a cargo de las sales biliares, lo que facilita la digestión.
- Esta emulsión se ve favorecida por los
movimientos peristálticos intestinales
- Sales biliares
- Sintetizadas en el
hígado, y almacenadas
en la vesícula biliar
- Sintetizadas en el
hígado, y almacenadas
en la vesícula biliar
- Transforma
- Grandes gotas
lipídicas de la dieta
- a
- Gotitas (micelas)
- Facilitando la actuación
de las enzimas digestivas
- Facilitando la actuación
de las enzimas digestivas
- Gotitas (micelas)
- a
- Grandes gotas
lipídicas de la dieta
- Esta emulsión se ve favorecida por los
movimientos peristálticos intestinales
- Para que se produzca la correcta asimilación de los lípidos, deben ser
hidrolizados por las enzimas digestivas hasta formar compuestos antipáticos,
que si puede atravesar las membranas, principalmente a nivel del yeyuno.
- Los lípidos no pueden atravesar libremente las membranas de
las células epiteliales del intestino ni enterocitos, debido a la
presencia de una capa de agua que rodea las microvellosidades.
- Enzimas digestivas
- Sobre
- Triacilglicéridos
- Actúan
- Lipasa pancreática
- Resultado
- Resultado
- Colipasa
- Resultado
- Un monoacilglicerol
- Sustancias antipáticas fáciles de
asimilar por las células de la
mucosa intestinal.
- Sustancias antipáticas fáciles de
asimilar por las células de la
mucosa intestinal.
- Dos moléculas de ácidos grasos
- En algunos casos también glicerol
- Sustancias antipáticas fáciles de
asimilar por las células de la
mucosa intestinal.
- Sustancias antipáticas fáciles de
asimilar por las células de la
mucosa intestinal.
- Un monoacilglicerol
- Resultado
- Lipasa pancreática
- Actúan
- Fosfolípidos
- Actúa
- Fosfolipasa A2
- Resultado
- Un ácido graso
- Asimilados por los enterocitos
- Asimilados por los enterocitos
- Un acil lisofosfolípido
- Asimilados por los enterocitos
- Asimilados por los enterocitos
- Un ácido graso
- Resultado
- Fosfolipasa A2
- Actúa
- Ésteres de colesterol
- Actúa
- Colesterol esterasa
- Resultado
- Colesterol
- Asimilados por los enterocitos
- Asimilados por los enterocitos
- Ácidos grasos
- Asimilados por los enterocitos
- Ya en el interior, vuelven a su forma inical.
- Su transporte son las lipoproteínas
- Su transporte son las lipoproteínas
- Ya en el interior, vuelven a su forma inical.
- Asimilados por los enterocitos
- Colesterol
- Resultado
- Colesterol esterasa
- Actúa
- Triacilglicéridos
- Sobre
- Emulsión de las
grasas
ingeridas
- Lipoproteínas
- Transportan los lípidos por el organismo
a nivel sanguíneo y linfáticos.
- Colaboran en el transporte de
aminoácidos
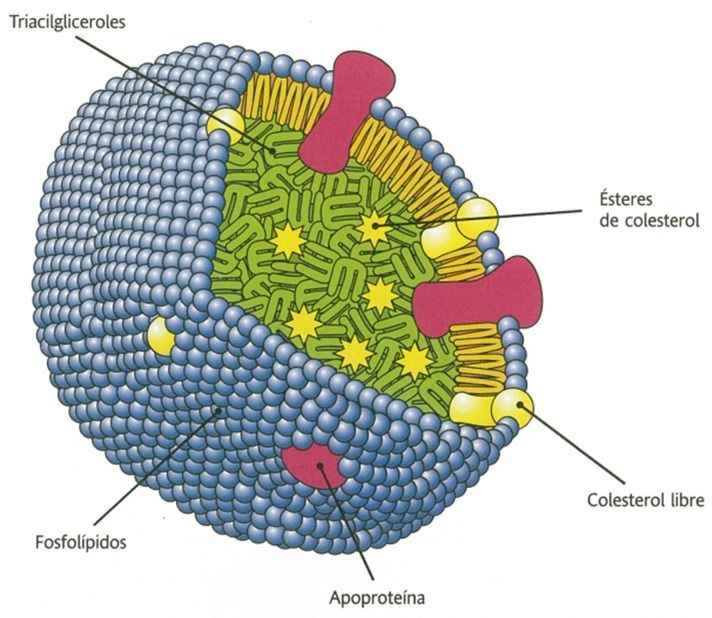
- Estructuras
complejas
- Estructura
- Capa externa
- Componentes
- Colesterol libre
- De naturaleza anfipática
- De naturaleza anfipática
- Apoproteínas
- De naturaleza anfipática
- De naturaleza anfipática
- Fosfolípidos
- De naturaleza anfipática
- De naturaleza anfipática
- Colesterol libre
- Componentes
- Interior
- Componentes
- Triacilglicéridos
- Hidrofóbicos
- Hidrofóbicos
- Colesterol
esterificado
- Hidrofóbicos
- Hidrofóbicos
- Triacilglicéridos
- Componentes
- Capa externa
- Principal
lipoproteína del
intestino
- Quilomicrón
- Transportados
hasta la sangre
- Llegan primero a los
tejidos periféricos y
luego al hígado
- De este modo, facilita la
acumulación de grasas en el
músculo y tejido adiposo
- De este modo, facilita la
acumulación de grasas en el
músculo y tejido adiposo
- Transportados
hasta la sangre
- Quilomicrón
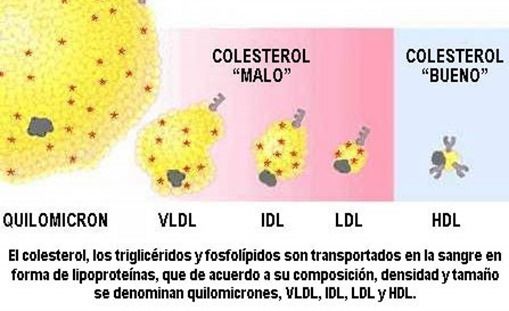
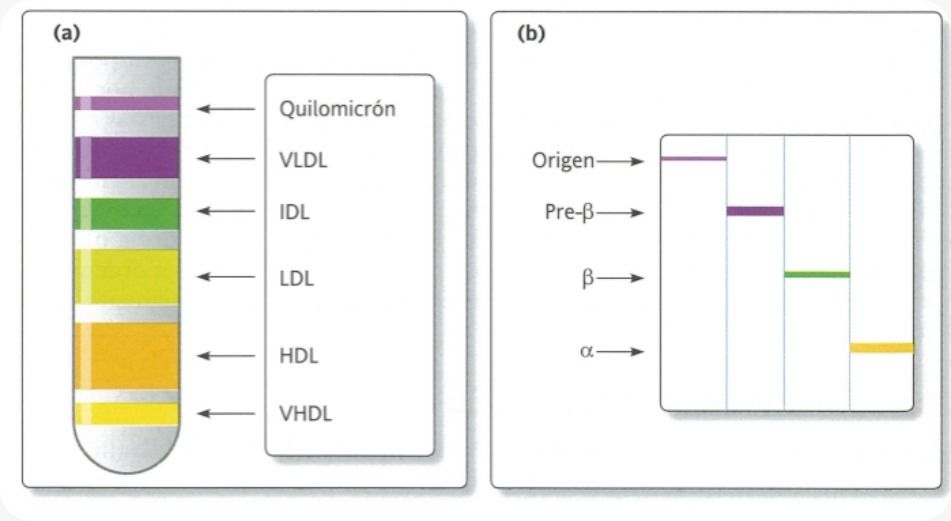
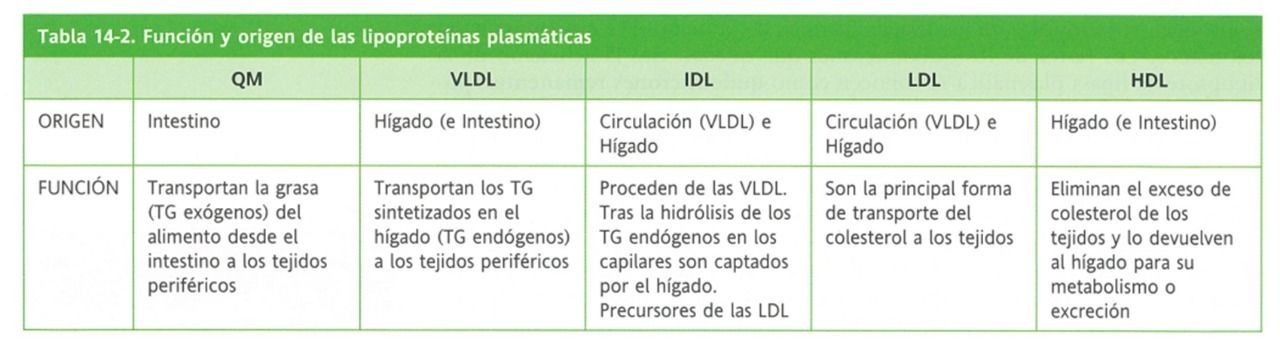
- Tipos
- Quilomicrones
(QM)
- Transportan los
triacilglicéridos exógenos
(de la dieta a los tejidos)
- Quilomicrones
remanentes
- Loss que quedan tras la actuación de la enzima.
(Contienen fosfolípidos y apoproteínas), que
son aprovechados por el hígado
- Loss que quedan tras la actuación de la enzima.
(Contienen fosfolípidos y apoproteínas), que
son aprovechados por el hígado
- Transportan los
triacilglicéridos exógenos
(de la dieta a los tejidos)
- Las diferencias de densidad permiten su
aislamiento mediante técnicas de
ultracentrifugación o de electrofóresis.
- A medida que el porcentaje de triacilglicéridos
disminuye, va aumentando el porcentaje de
proteínas y colesterol libre o esterificado Esto
determina que la densidad vaya en aumento.
- VLDL
- De muy baja
densidad
- Transportan los triacilglicéridos
endógenos (propios del organismo,
sintetizados a nivel hepático)
- De muy baja
densidad
- IDL
- De densidad
intermedia
- De densidad
intermedia
- LDL
- De densidad
baja
- Fuente de colesterol
para los tejidos
periféricos, también
aportan fosfolípidos y
aminoácidos.
- De densidad
baja
- HDL
- De densidad
alta
- Recogen el exceso de colesterol depositado en
los tejidos periféricos y los retiran al hígado
- De densidad
alta
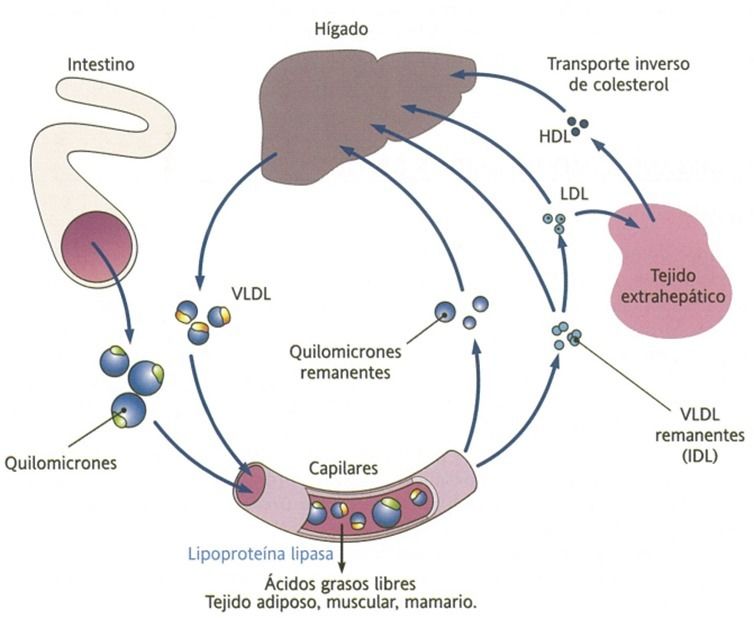
- Quilomicrones
(QM)
- En el intestino, además de la
formación de quilomicrones,
también se puede originar VLDL.
- Estas se vierten en la linfa hasta la
sangre, al no pasar por la circulación
enterohepática, llegan primero a los
tejidos periféricos.
- En estos, la enzima lipoproteína lipasa plasmática
hidroliza a los triacilglicéridos en glicerol y
ácidos grasos, asimilados por las células tisulares
(adipocitos y miocitos).
- El glicerol y ácidos grasos entran por difusión
simple a los tejidos periféricos, forman
triacilglicéridos y almacenan grandes
cantidades de energía cuándo sea necesaria.
- El glicerol y ácidos grasos entran por difusión
simple a los tejidos periféricos, forman
triacilglicéridos y almacenan grandes
cantidades de energía cuándo sea necesaria.
- En estos, la enzima lipoproteína lipasa plasmática
hidroliza a los triacilglicéridos en glicerol y
ácidos grasos, asimilados por las células tisulares
(adipocitos y miocitos).
- Estas se vierten en la linfa hasta la
sangre, al no pasar por la circulación
enterohepática, llegan primero a los
tejidos periféricos.
- La actuación de la lipoproteína lipasa sobre las VLDL
originan glicerol, ácidos grasos, IDL o VLDL
remanentes (ricas en colesterol)
- Estas pueden enriquecerse de colesterol gracias a la CETP
(proteína transportadora de colesterol esterificado), la cual
provoca que las IDL se transformen en LDL.
- Las LDL son retiradas por las células
de los tejidos periféricos a través de
transportadores específicos
- A nivel sanguíneo, aparecen las HDL, de origen
hepático y que reciben el exceso de colesterol
depositado en los tejidos periféricos y
transportado al hígado.
- La enzima LCAT (lecitina colesterol
aciltransferasa) esterifica el
colesterol recogido por las HDL.
- La enzima LCAT (lecitina colesterol
aciltransferasa) esterifica el
colesterol recogido por las HDL.
- A nivel sanguíneo, aparecen las HDL, de origen
hepático y que reciben el exceso de colesterol
depositado en los tejidos periféricos y
transportado al hígado.
- Las LDL son retiradas por las células
de los tejidos periféricos a través de
transportadores específicos
- Estas pueden enriquecerse de colesterol gracias a la CETP
(proteína transportadora de colesterol esterificado), la cual
provoca que las IDL se transformen en LDL.
- Transportan los lípidos por el organismo
a nivel sanguíneo y linfáticos.
- Metabolismo de
ácidos grasos
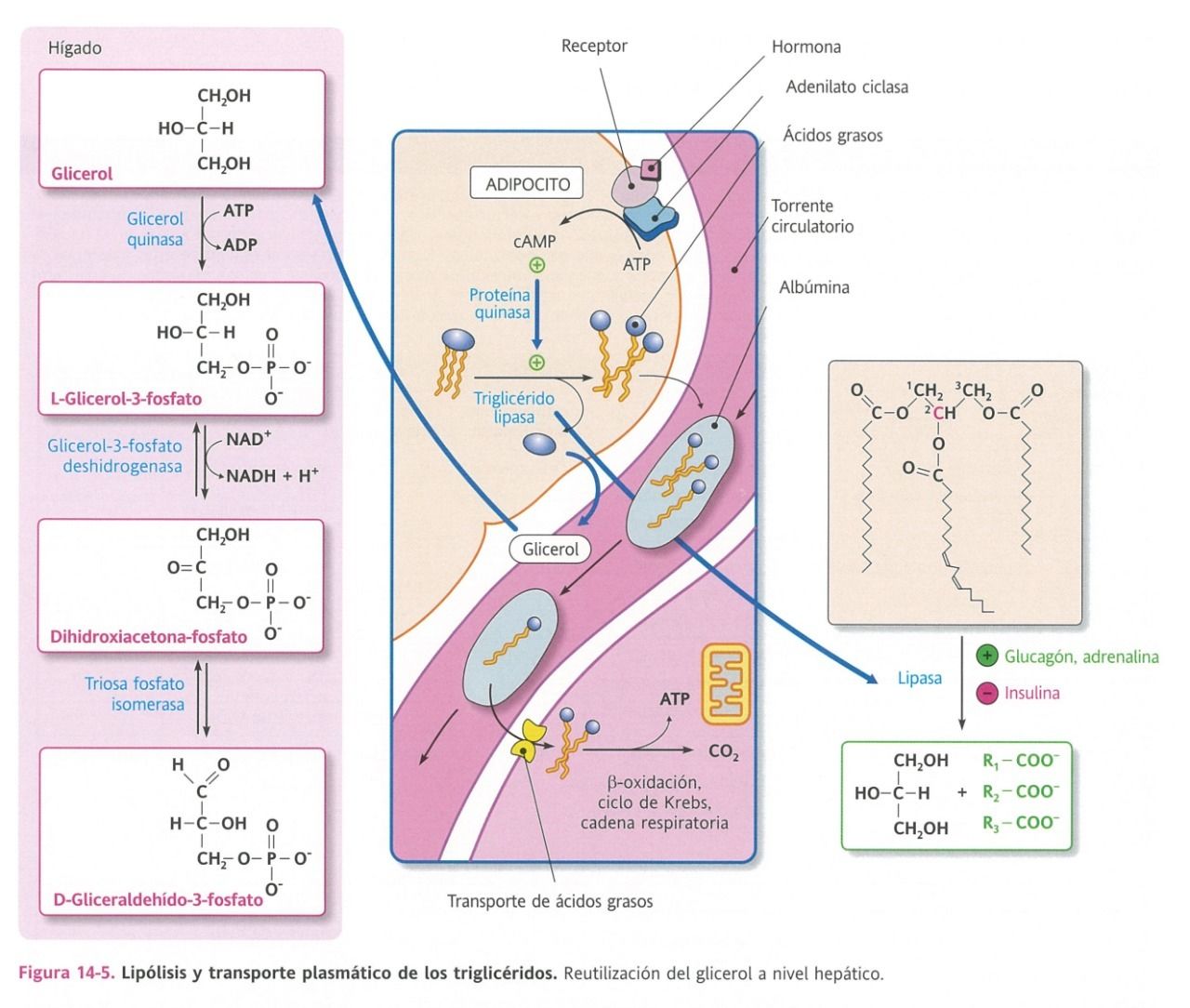
- Lipólisis
- Mecanismo de movilización de
los lípidos almacenados como
reservorio de energía
- Estos lípidos acumulados se encuentran como gotitas
(anhidra) en el citoplasma de las células adiposas.
- Estos lípidos acumulados se encuentran como gotitas
(anhidra) en el citoplasma de las células adiposas.
- Sucede cuando hay una
deficiencia del aporte
energético o cuando se ayuna
- Hidrólisis
- Primer paso de
su catabolismo
- Triglicérido
lipasa
intracelular
- Resultado
- Glicerol
- 3 ácidos
grasos
- Glicerol
- Esta enzima es potenciada por el glucagón y la
adrenalina, favoreciendo la lipólisis, al fosforilar a
la triacilglicérido lipasa a través de la proteína
quinasa A dependiente de AMP.
- Mientras que la insulina, al potenciar una fosfatasa que
desfosforila la lipoproteína lipasa, bloquea la lipólisis.
- Resultado
- Actúa
- Los ácidos grasos salen del adipocito y se unen en sangre a la
albumina (VHDL), la cual puede transportar entre dos, cuatro
hasta 6 ácidos grasos.
- Estos llegan hasta los tejidos
que requieren energía
- Hígado
- Músculo cardíaco
- Músculo esquelético
- En estos los ácidos grasos son oxidados
en la vía metabólica beta-oxidación,
para producir energía.
- Hígado
- Estos llegan hasta los tejidos
que requieren energía
- El glicerol también
sale a la sangre
- Se transforma en
- Dihidroxiacetona
- En el hígado
- Entra a la gluconeogénesis a
nivel hepático o a la vía
glucolítica para producir energía.
- En el hígado
- Dihidroxiacetona
- Se transforma en
- Primer paso de
su catabolismo
- Mecanismo de movilización de
los lípidos almacenados como
reservorio de energía
- Degradación de
ácidos grasos
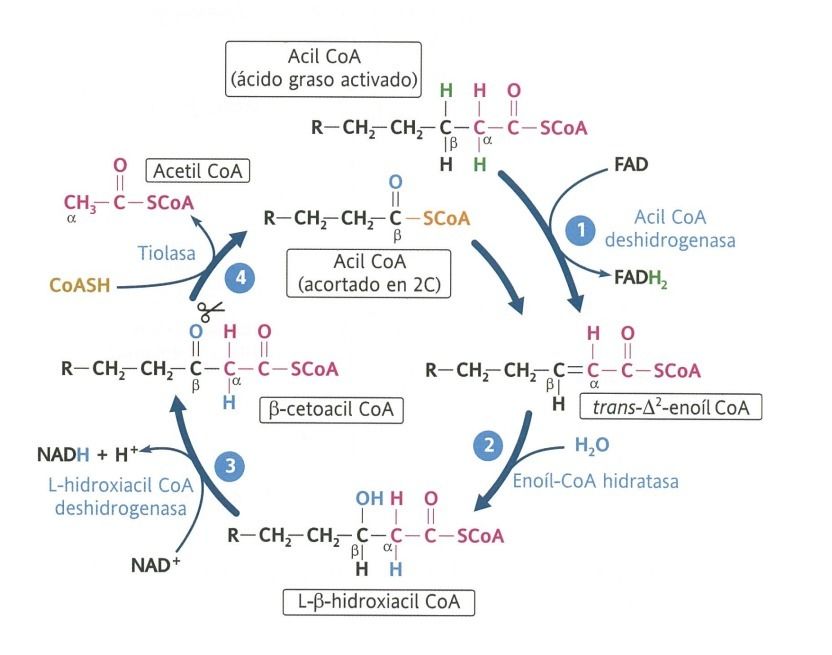
- Se producen sucesivas oxidación en el carbono beta, que
van separando fragmentos de dos carbonos en forma de
acetileno CoA, que se incorporarán al ciclo de Krebs.
- Implica
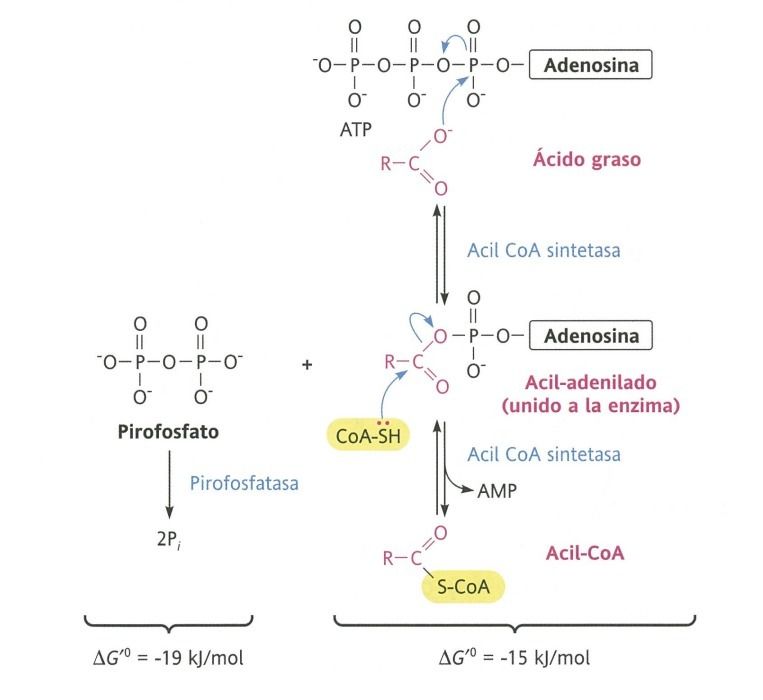
- Fase 1
- La activación del ácido graso esterificándose
con el CoA y a expensas del ATP
- Esta activación la realiza la acil CoA sintetasa en la
membrana externa de la mitocondria.
- La actuación de esta forma
las moléculas de acil CoA
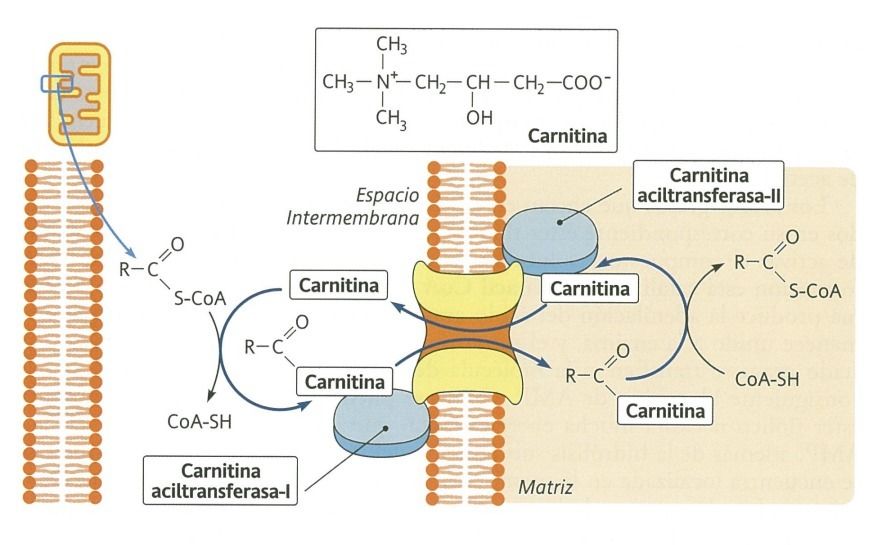
- Al ser muy largas necesitan ayuda para ingresar a la
mitocondria, a través de un sistema de lanzadera
- En el que intervienen
- Carnitina
- Carnitina acil tranferasa-I
- Carnitina acil-carnitina translocasa
- Carnitina acil transferasa-II
- Carnitina
- Una vez dentro de la matriz mitocondrial, las
moléculas de acil CoA comienzan la
degradación propia de la beta-oxidación.
- 2. Hidratación
- 1. Deshidrogenación
- 3. Deshidrogenación
- 4. Ruptura tiólica
- 2. Hidratación
- En el que intervienen
- Al ser muy largas necesitan ayuda para ingresar a la
mitocondria, a través de un sistema de lanzadera
- La actuación de esta forma
las moléculas de acil CoA
- Esta activación la realiza la acil CoA sintetasa en la
membrana externa de la mitocondria.
- La activación del ácido graso esterificándose
con el CoA y a expensas del ATP
- Fase 2
- Entrada a la mitocondria por un
transporte mediado por la carnitina
- Entrada a la mitocondria por un
transporte mediado por la carnitina
- Fase 3
- Beta-oxidación propiamente, degradándose el
ácido graso a moléculas de acetil CoA.
- Beta-oxidación propiamente, degradándose el
ácido graso a moléculas de acetil CoA.
- Fase 1
- Se producen sucesivas oxidación en el carbono beta, que
van separando fragmentos de dos carbonos en forma de
acetileno CoA, que se incorporarán al ciclo de Krebs.
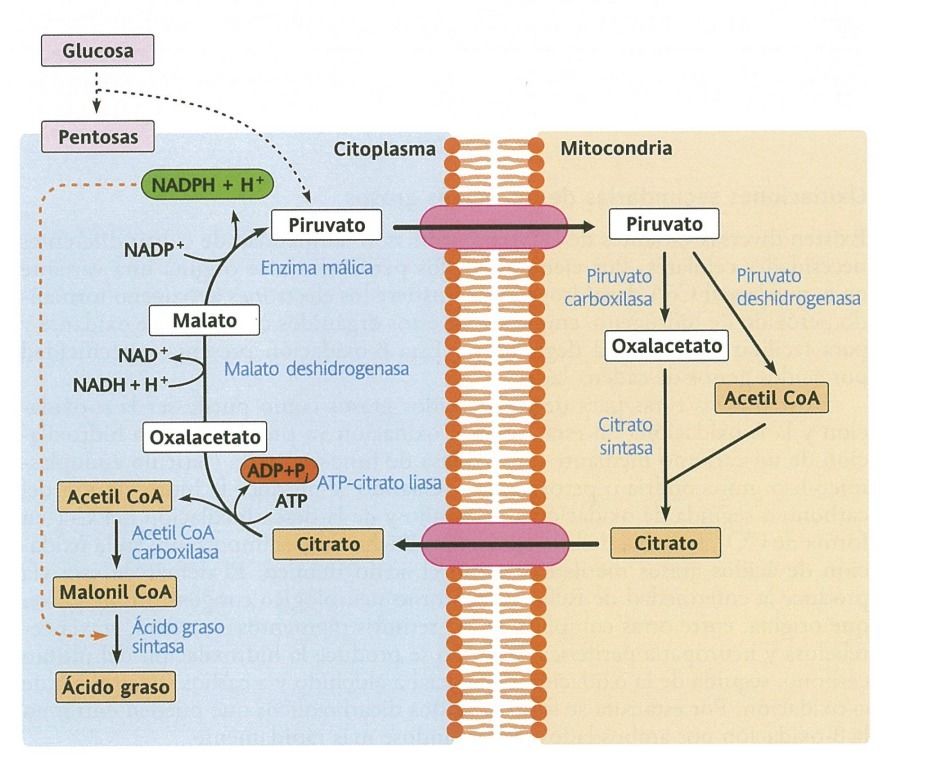
- Biosintesis de
ácidos grasos
- Debido a la limitada capacidad para almacenar glucosa,
la ruta biosintética se convierte en una vía importante.
- La glucosa ingerida en exceso se convierte en ácidos
grasos y estos en triacilglicéridos que pueden almacenarse
en grandes cantidades en el tejido adiposo.
- Está síntesis es posible incluso en citoplasma sin
mitocondrias, e incluso aumenta su capacidad en estos casos.
- Está síntesis es posible incluso en citoplasma sin
mitocondrias, e incluso aumenta su capacidad en estos casos.
- Para proceder a la síntesis de ácidos grasos se requiere poder
reductor (NADPH + H+) y moléculas de malonil CoA.
- Está regulada principalmente a nivel hormonal, siendo
favorecida por la insulina e inhibida por el glucagón.
- Debido a la limitada capacidad para almacenar glucosa,
la ruta biosintética se convierte en una vía importante.
- Lipólisis
- Cuerpos cetónicos
- Sustancias que se producen a partir de CoA en las mitocondrias del tejido
hepático cuando la velocidad de la beta-oxidación supera a la velocidad de la
oxidación del acetil CoA en el ciclo de Krebs (situaciones de ayuno).
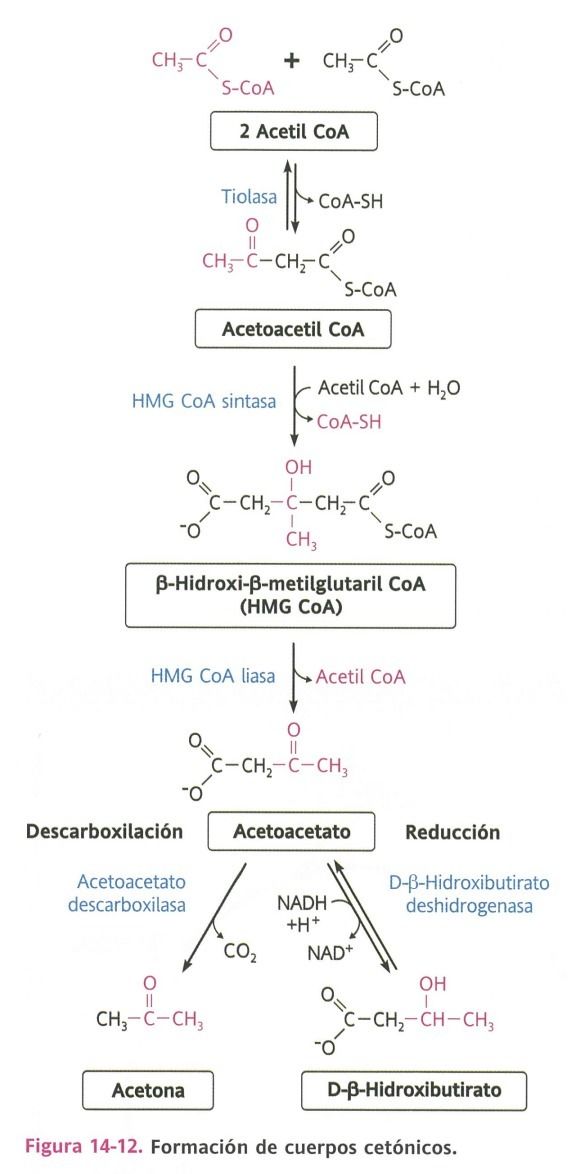
- Acetoacetato
- Hidroxibutirato
- Acetona
- Acetoacetato
- Estos sirven como fuente de energía para
el corazón, el músculo y otros tejidos.
- Favorecen un ahorro
de glucosa
- Si se produce un ayuno muy prolongado pueden ser utilizados por el
cerebro como fuente de energía alternativa a la glucosa.
- Estos compuestos se utilizan ya que los animales no puede transformar de forma neta
los ácidos grasos en hidratos de carbono, al carecer del ciclo del glioxilato.
- Cetogénesis
- Proceso de creación de los
cuerpos cetónicos.
- Condensación de dos moléculas de
acetil CoA por acción de una tiolasa,
formando el acetoacetil CoA.
- Luego se fusiona con
un acetil CoA
- Originando
- Hidroximetilglutaril CoA.
- Sirve para la síntesis de los cuerpos cetónicos y la biosíntesis del colesterol
- Sirve para la síntesis de los cuerpos cetónicos y la biosíntesis del colesterol
- Hidroximetilglutaril CoA.
- Originando
- Luego se fusiona con
un acetil CoA
- Proceso de creación de los
cuerpos cetónicos.
- Utilización
- Los cuerpos cetónicos asimilados por los tejidos extrahepáticos se utilizan para
producir moléculas de acetil CoA que se degradarán en el ciclo de Krebs.
- El uso de los cuerpos cetónicos depende de las fluctuaciones de los niveles de glucosa en sangre.
- Se usa especialmente en períodos prolongados de ayuno.
- Se usa especialmente en períodos prolongados de ayuno.
- Los cuerpos cetónicos asimilados por los tejidos extrahepáticos se utilizan para
producir moléculas de acetil CoA que se degradarán en el ciclo de Krebs.
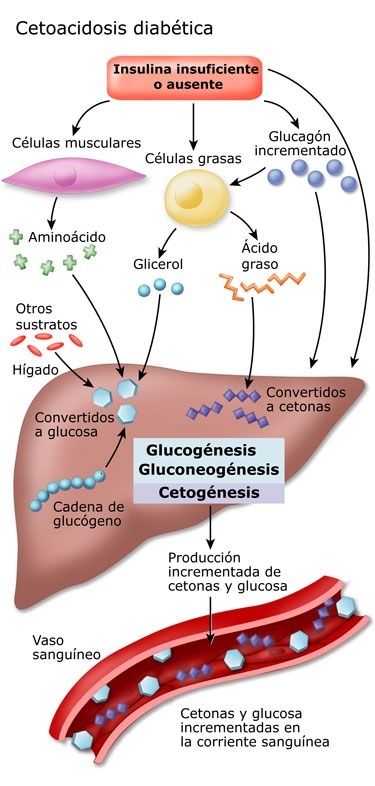
- Cetoacidosis
- Causado por
- Un incremento importante de los niveles de los cuerpos cetónicos en sangre
- Un incremento importante de los niveles de los cuerpos cetónicos en sangre
- Trastorno grave que puede ser mortal.
- La más conocida es la cetoacidosis diabética, causado por una deficiencia absoluta o relativa de insulina.
- Causado por
- Sustancias que se producen a partir de CoA en las mitocondrias del tejido
hepático cuando la velocidad de la beta-oxidación supera a la velocidad de la
oxidación del acetil CoA en el ciclo de Krebs (situaciones de ayuno).
- Biosíntesis de lípidos
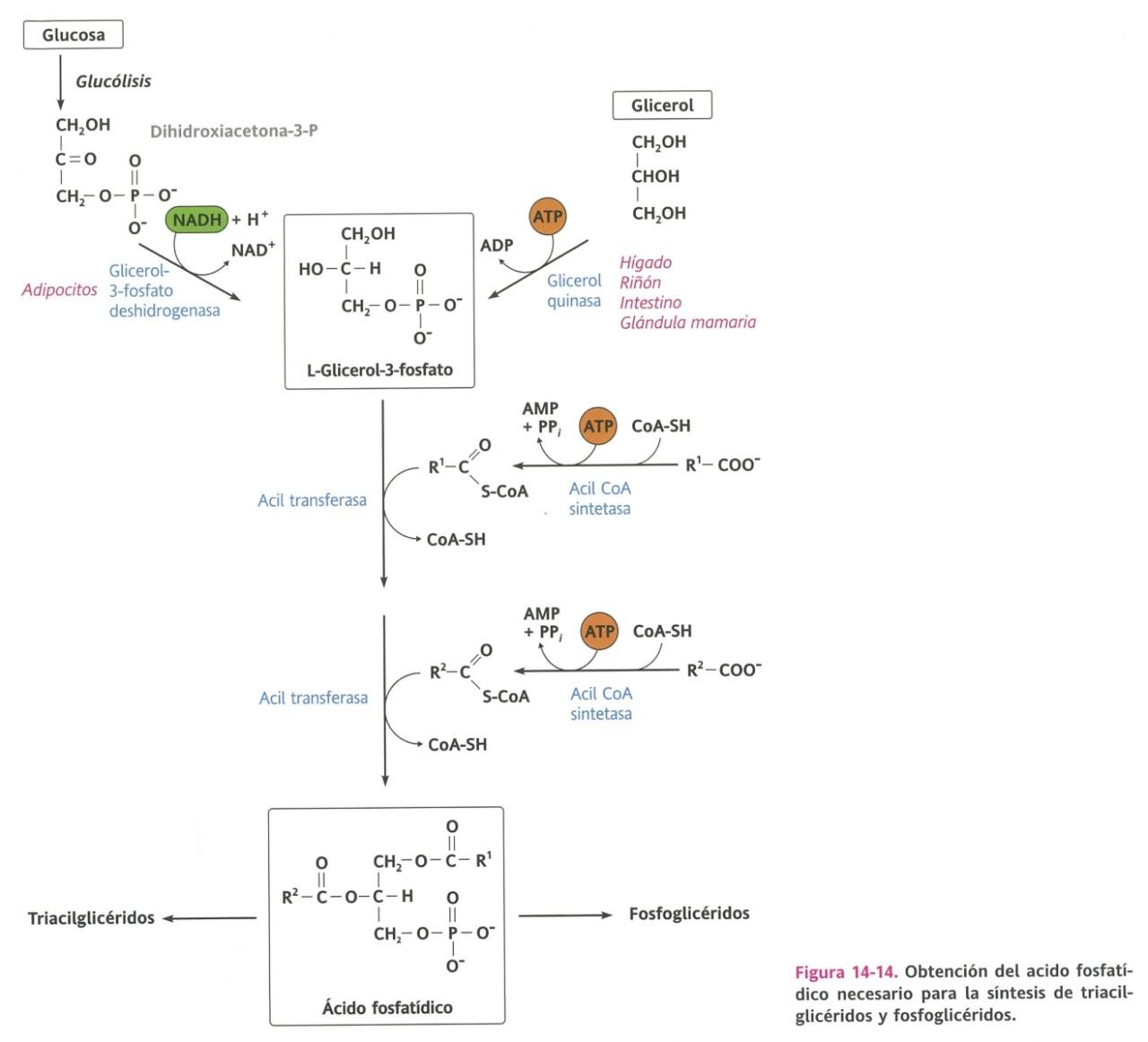
- Biosíntesis de
acilglicéridos
- La síntesis de los triacilglicéridos tiene lugar en el retículo
endoplasmático liso (REL) de células adiposas y hepáticas.
- Se origina mediante la esterificación secuencial de una molécula de
glicerol-3-fosfato con tres moléculas de acil CoA (ácidos grasos activados).
- Requiere de un fosfolípido
intermediario:
- Ácido
fosfatídico
- Síntesis del Ácido
fosfatídico
- 1. Síntesis de
glicerol-3-fosfato
- 2. Activación de los
ácidos grasos
- 3. Transferencia de los
ácidos grasos activados
- 1. Síntesis de
glicerol-3-fosfato
- Participa en la síntesis de
triacilglicéridos y de
fosfoglicéridos.
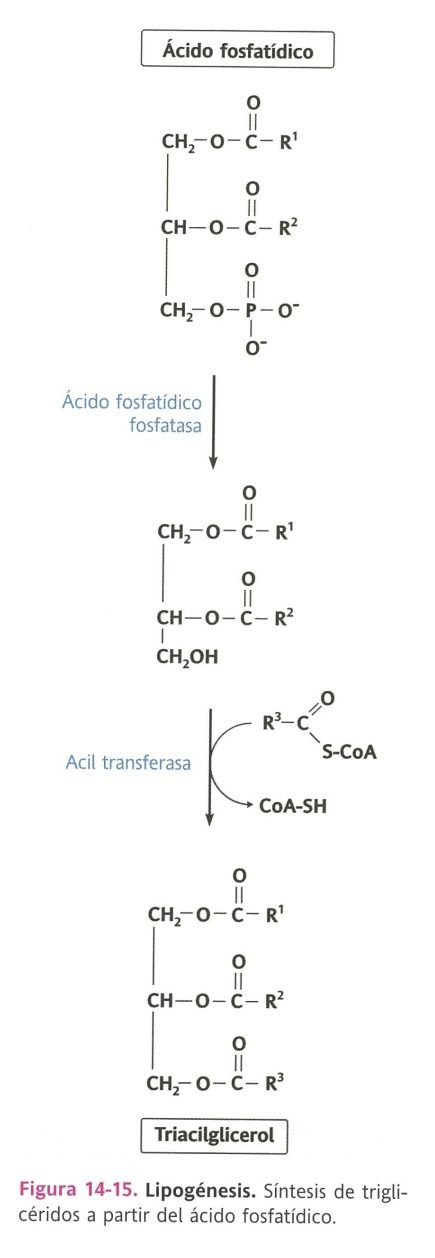
- En la síntesis de triacilglicéridos, éste ácido debe desprenderse del grupo fosfato de la posición 3,
gracias a la fosfatasa, esto resulta en un diacilglicerol, el cual se transformará en triacilglicerol
mediante la acil tranferasa que transferirá un ácido graso a la posición 3.
- Los triacilgliceroles se almacenan en el citoplasma o incorporarse en
lipoproteínas, intestino delgado e hígado, o leche en la glándula mamaria.
- Los triacilgliceroles se almacenan en el citoplasma o incorporarse en
lipoproteínas, intestino delgado e hígado, o leche en la glándula mamaria.
- En la síntesis de triacilglicéridos, éste ácido debe desprenderse del grupo fosfato de la posición 3,
gracias a la fosfatasa, esto resulta en un diacilglicerol, el cual se transformará en triacilglicerol
mediante la acil tranferasa que transferirá un ácido graso a la posición 3.
- Síntesis del Ácido
fosfatídico
- Ácido
fosfatídico
- Requiere de un fosfolípido
intermediario:
- Se origina mediante la esterificación secuencial de una molécula de
glicerol-3-fosfato con tres moléculas de acil CoA (ácidos grasos activados).
- La síntesis de los triacilglicéridos tiene lugar en el retículo
endoplasmático liso (REL) de células adiposas y hepáticas.
- Biosíntesis del colesterol
- La HMG CoA reductasa es la enzima clave de la
síntesis de colesterol, esta fuertemente regulada y es
una importante diana farmacológica.
- En los vertebrados, la síntesis del colesterol está controlado
principalmente mediante la velocidad a la que el colesterol
ingresa en las células procedente del torrente sanguíneo.
- Tiene lugar en el citoplasma a partir de las
moléculas de acetil CoA.
- Etapas
- Tercera
etapa:
- Ciclación del escualeno a
lanosterol (C30) y
conversión final a
colesterol (C27).
- Ciclación del escualeno a
lanosterol (C30) y
conversión final a
colesterol (C27).
- Segunda
etapa:
- Condensación de seis moléculas
de isoprenos activados para
formar escualeno.
- Condensación de seis moléculas
de isoprenos activados para
formar escualeno.
- Primera
etapa:
- Síntesis de los
isoprenos activados a
partir de acetil CoA.
- Síntesis de los
isoprenos activados a
partir de acetil CoA.
- Tercera
etapa:
- Etapas
- La HMG CoA reductasa es la enzima clave de la
síntesis de colesterol, esta fuertemente regulada y es
una importante diana farmacológica.
- Biosíntesis de
acilglicéridos
Media attachments
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Want to create your own Mind Maps for free with GoConqr? Learn more.