18154206
6. RNA Processing
Descrição
Sem etiquetas
Slides por Chloe Cavarretta, atualizado more than 1 year ago
Mais
Menos
|
|
Criado por Chloe Cavarretta
mais de 6 anos atrás
|
|
Resumo de Recurso
Slide 1
Learning Outcomes: L13
Describe how the 5’ and 3’ ends of most eukaryotic mRNAs are formed
Understand the basic mechanisms of splicing and how splicing regulatory proteins can influence how splice sites are selected
Understand that alternative splicing can increase the coding capacity of genomes
Solve problems based on your understanding of splicing
Slide 2
Overall Production of Mature mRNA
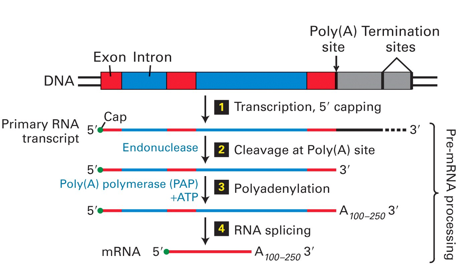{kind=link}
Slide 3
Capping on 5' End of Transcripts
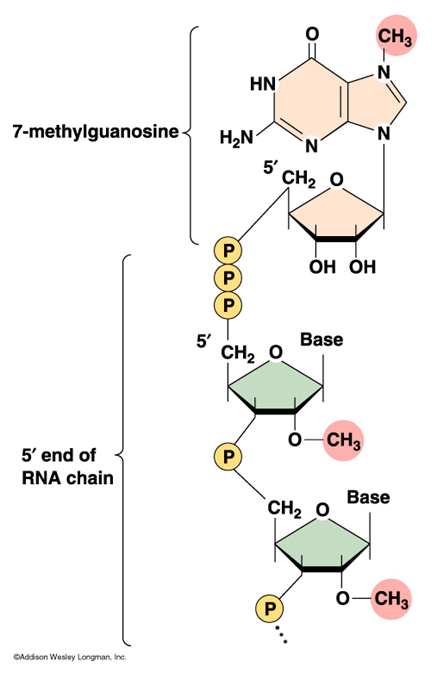{kind=link}
3 enzymes act in succession:
Phosphatase- removes phosphate from original 5' end of mRNA
Guanyl transferase- adds guanosine base at 5' end
Methyltransferase- guanosine gets methylated
Usually phosphodiester backbone is between 5' and 3' carbons but cap causes linkage of 5' to 5' carbons
Avoids cap getting broken by enzymes that digest 5' to 3' linkages
Slide 4
Capping on 5' End of Transcripts
Capping occurs co-transcriptionally
Newly formed pre-mRNA is capped early
Capping enzymes bind phosphorylated RNAP II CTD as it transcribes (ride along)
When transcript emerges from RNAP II (25-30bp) enzymes add the modification, RNAP pauses whilst modifying and will continue when finished
Slide 5
Poly-A Tail at 3' End of Transcripts
Cleavage and Polyadenylation
The 3' end of mRNA is shorter than the original transcript - cleaved to add the polyA tail on (upto 200 adenosine residues)
Need a consensus sequence for cleavage and polyadenylation events- mainly CA dinucleotide
CA cleavage site is flanked with other consensus sequences upstream and downstream where proteins can bind
When cleavage occurs the 3' end is degraded in the nucleus and get addition of polyA
Slide 6
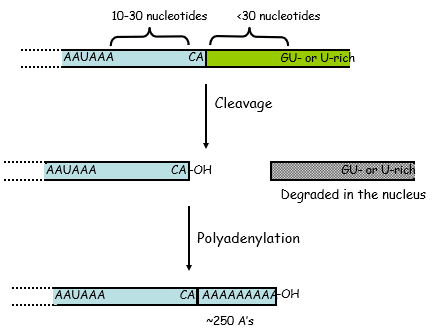{kind=link}
Poly-A Tail at 3' End of Transcripts
Slide 7
Proteins Involved in Cleavage/Adding PolyA
Proteins recognise and bind the flanking sequences:
Cleavage stimulation factor F (CstF)
Cleavage and polyadenylation specificity factor (CPSF)
These proteins also ride along the phosphorylated CTD tail of RNAP II during transcription
When the sequences they bind are transcribed they transfer to the 3' end sequences on mRNA
The proteins then bind each other and present CA nucleotide cleavage site to cleavage factors
Slide 8
Proteins Involved in Cleavage/Adding PolyA
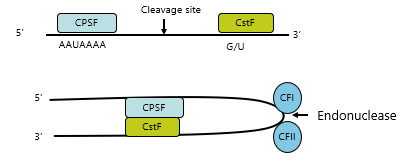{kind=link}
Slide 9
Following Cleavage
An enzyme poly-A-polymerase (PAP) sequentially adds ~200 A nucleotides (from ATP precursor)
Poly-A binding proteins (PABP) assemble onto the PolyA tail
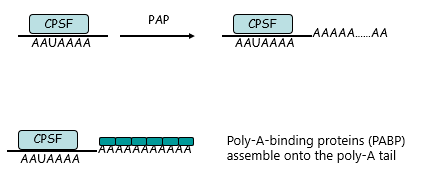{kind=link}
Slide 10
Why Add 5' Cap and 3' PolyA Tail?
Stabilisation:
Cap, PolyA tail and PolyA binding proteins promote stabilisation
PolyA proteins bind to other proteins that bind to Cap= circulisation of mRNA which protects ends being chewed by enzymes
Leave nucleus into cytoplasm which needs stabilised mRNA
Involvement in export:
mRNA needs to be transported to cytoplasm to be translated
Proteins that do shuttling of RNAs out of nucleus recognise cap and polyA/(PABPs)
Involvement in translation:
Translation is initiated through cap binding proteins that recognise cap
Slide 11
RNA Splicing
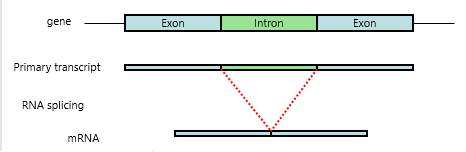{kind=link}
Need to accurately remove introns as single base errors will shift reading frames
Introns aren't junk:
Allow alternative splicing
Promote gene expression
Promote export out of the nucleus
Slide 12
RNA Splicing: Sequence Signal for Splice Site
Signal to guide machinery to point of splicing
High conservation of sequence is found only immediately within intron at junctions (at exon-intron boundary)
Generally GU as first base (5' splice site)
Often run of pyrimidines then AG (3' splice site)
A= branchpoint site
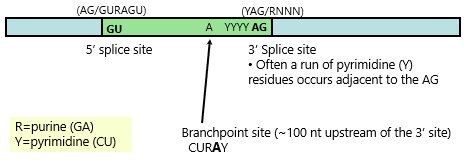{kind=link}
Slide 13
RNA Splicing: Mechanism
Nucleophilic attack of 2'OH from A onto exon 1 (5' splice site)
Releases exon 1 and forms loop back on itself
Nucleophillic attack of 3' OH from exon 1 onto exon 2 (3' splice site)
Exons join together and excising of lariat structure
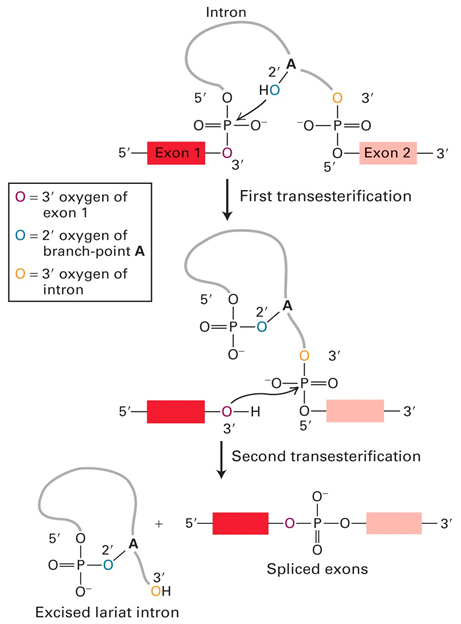{kind=link}
Slide 14
Spliceosome
Performs splicing
A highly complex molecular machine composed of small nuclear ribonucleoprotein particles (snRNPs/snurps)
snRNPs made of U RNAs and proteins
snRNPs find the spice site boundaries to ensure reactions happen in the right place
5 U RNAs in total and multiple proteins involved
Consensus sequences recognised by the RNA component, each snRNP has 1 RNA and lots of proteins
Slide 15
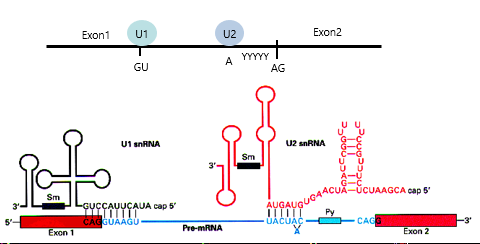
Spliceosome Steps of Splicing
U1 snRNP binds 5' splice site and U2 snRNP binds branch point
RNA pairing is crucial in bringing the spliceosome to the correct place in the pre-mRNA
U1 snRNA pairs with pre-mRNA at exon intron boundary of 5' splice site
U2 snRNA pairs at branch point
Not perfectly paired- binding of U2 to branch point forces A out so it is unpaired and activated for nucleophilic attack on 5' splice site
Slide 16
Spliceosome Steps of Splicing
{kind=link}
Slide 17
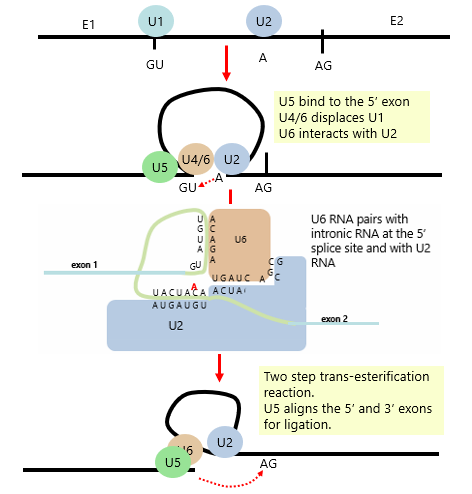
Spliceosome Steps of Splicing
U1 is displaced by U4/6, U6 interacts with U2, U5 binds the 5' exon sequence
U6 RNA pair with intronic RNA at 5' splice site and with U2 RNA
Pairing of U2 and U6 ensures A is at the exact right position of the 5' side of GU (5' splice site) for nucelophilic attack onto exon 1
Two step trans-esterification (2 Nu attacks) and U5 aligns the 5' and 3' exons for ligation
Slide 18
{kind=link}
Spliceosome Steps of Splicing
Slide 19
Challenges of Splice Site Identification
Most genes have multiple introns- need correct exons to join
Adv that co-transcriptionally as not all introns out at one time
Many splice site consensus sequences are not perfectly conserved
Requires RNA-RNA interactions, weak splice sites have weak interactions between RNA-RNA (less complementary)
So spliceosome needs more help, there are other sequences in mRNA that can be recognised by RNA binding proteins and help recruit the spliceosome
Slide 20
Other Sequences that Recruite Spliceosome
Exonic splicing enhancers (ESEs) interact with SR proteins (conserved family of Serine/Arginine rich splicing factors)
Single base change in ESE can affect binding of SR proteins, therefore effect whether splicing occurs
SR proteins recognise sequences within exons (ESEs) and recruit and stabilise splicing machinery via protein-protein interactions
SR proteins regulated (eg phosphorylation) to activate/deactivate
Way of achieving alternative splicing
Slide 21
Alternative Splicing
Increase protein coding capacity of higher eukaryotes
But most genes have a major isoform and the proteome is smaller than that predicted from the transcriptome
Proteome= entire set of proteins expressed in a cell at a certain time
Transcriptome= entire set of RNAs expressed in a cell at certain time
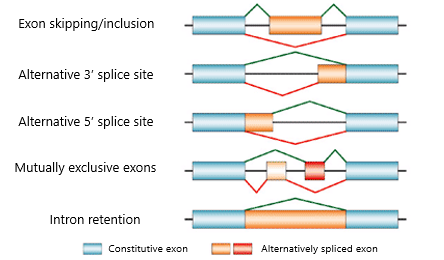
Impact of alternative RNA splicing:
In 5'UTR= altered translation, including removal of regulatory elements that impact translation
In protein coding sequence (ORF)= altered protein structure, produce different protein isoforms
In 3'UTR= altered mRNA stability
Slide 22
Patterns of Alternative Splicing
{kind=link}
Slide 23
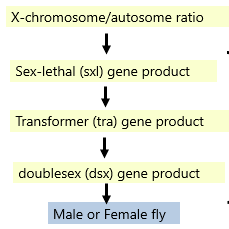
Example of Effect of Alternative Splicing
The primary signal for determining whether fly develops as male or female is X chromosome/autosome ratio:
Female= 2X chromosomes and 2 sets of autosomes (ratio=1)
Male= 1X chromosome and 2 sets of autosomes (ratio = 0.5)
Cascade of alternative splicing events in 3 genes-->
{kind=link}
Slide 24
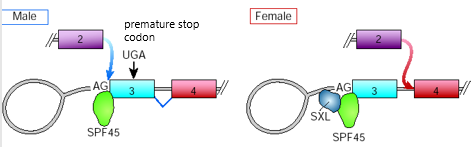
1. Sex-lethal (sxl) gene
Example of exon skipping/inclusion
SPF45 = splicing enhancer that promotes inclusion of exon 3
SXL = acting as an inhibitor of exon 3 inclusion
{kind=link}
Slide 25
Males:
Sxl transcript includes exon 3 that has premature stop codon
SPF45 binds AG dinucleotide and promotes exon 3 joining with exon 2
Because of pre-mature stop codon, produce non-functional sex lethal protein
Females:
SXL binds site adjacent SPF45 and interferes with its activity as a splicing enhancer
Exon 3 is skipped, exon 2 joins with exon 4
Produce functional sex lethal protein because 3' splice site of exon 3 not used
SXL regulates alternative splicing of its own pre-mRNA
Females
1. Sex-lethal (sxl) gene
Slide 26
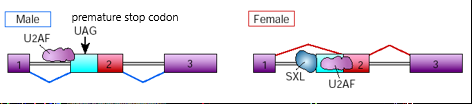
2. Transformer (tra) gene
Males:
Proximal 3' splice site used, promoted by U2AF auxiliary factor
mRNA produced contains premature stop codon= non-functional tra protein
Females:
SXL product binds pre-mRNA and shifts the binding of U2AF to a more distal 3' splice site of exon 2 - SXL acts as splice site repressor
Premature stop codon not included so functional tra protein formed
Slide 27
{kind=link}
2. Transformer (tra) gene
Slide 28
3. Doublesex (dsx) Gene
Male:
Dont have transformer product
3' splice site immediately upstream of exon 4 not recognised
Exon 4 skipped, use a more distal polyA site not on exon 4
Females:
Tra acts as promoter of exon inclusion by complexing with SR protein (RBP1) and SR-like protein (TRA2) to bind ESEs (sequence enhancers) in exon 4
Recruit splicing machinery to 3' splice site
Exon 4 has polyA signal which results in cleavage at that site, exon 5 and 6 not included
Slide 29
Summary of Example
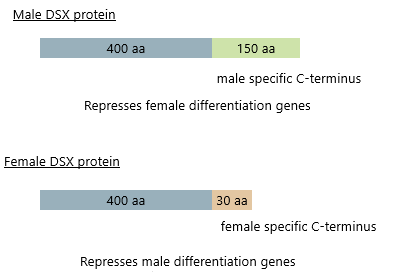{kind=link}
Both male and females produce variants of DSX protein- male protein represses female differentiation genes
Both have same N- because have same first 3 exons but have specific C-
Males have longer version
Controlled by proteins that recognise and block or recruit splicing activity
Quer criar seus próprios Slides gratuitos com a GoConqr? Saiba mais.