Description
|
|
Created by Cher Bachar
almost 13 years ago
|
|
Page 1
{kind=link}
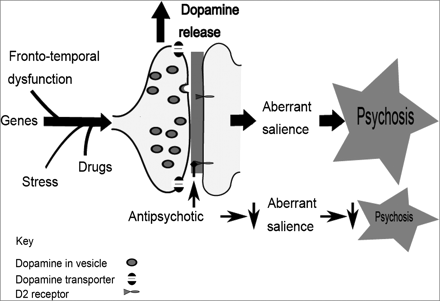
Multiple hits interact to result in striatal dopamine dysregulation to alter the appraisal of stimuli and resulting in psychosis, whilst current antipsychotic drugs act downstream of the primary dopaminergic dysregulation.
{kind=link}
{kind=link}
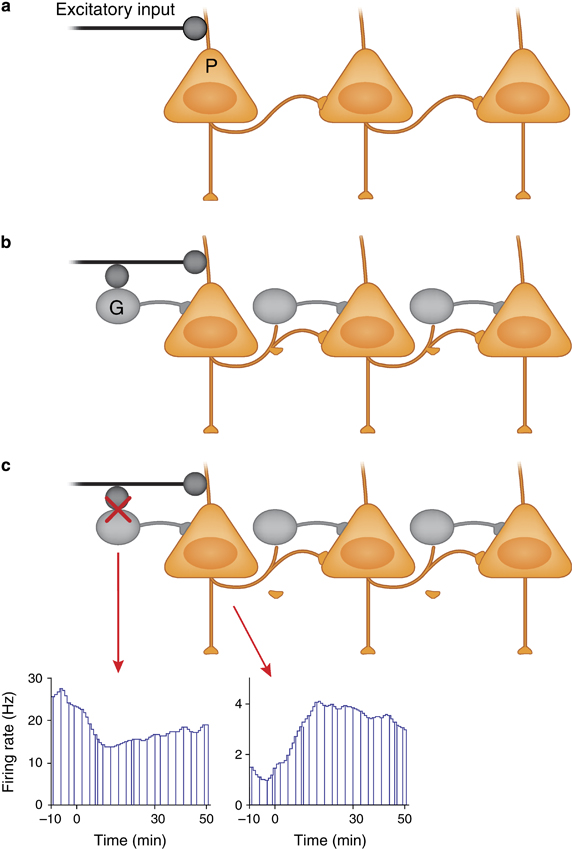
(a) Principal (P) or pyramidal cells in the neocortex and hippocampus, all which use glutamate as their neurotransmitter, receive extensive excitatory input from subcortical and cortical regions. In the absence of a counteracting inhibitory influence, activation of these inputs could cause a chain reaction of ever increasing excitation. (b) The regulation or stabilization of the firing of pyramidal cells is served by GABA (G) interneurons. One classic example of GABAergic influence is the feed-forward inhibition model where the effect of afferent excitation on a pyramidal neuron is dampened by co-activation of GABA interneurons that synapse onto the same pyramidal neuron. (c) The excitatory–inhibitory balance can be disrupted by many factors. An example is exposure to pro-psychotic compounds such as NMDA receptor antagonists. Blockade of NMDA receptors preferentially acts on fast spiking GABA interneurons because these neurons have a more depolarized membrane potential (note the higher firing rate on the recording from a putative GABA interneuron in the prefrontal cortex of an awake rat) and thus contain more open NMDA channels. This preferential inhibition of GABA interneurons creates an artificial state of disinhibition for the pyramidal cells and increases their firing rate.
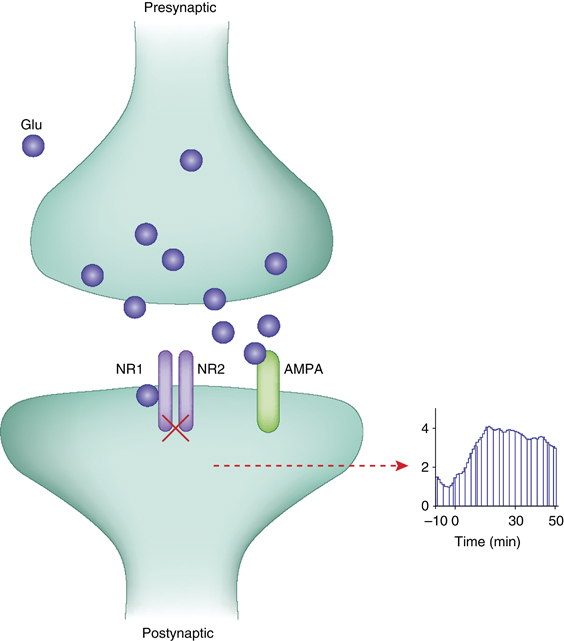
One of the downstream consequences of NMDAR inhibition is increased availability of glutamate (see text and Figure 2 for potential mechansims that can cause this effect). This increase causes excess activity of AMPA receptors and enhanced postsynaptic spiking of cortical principle cells at rest.
{kind=link}
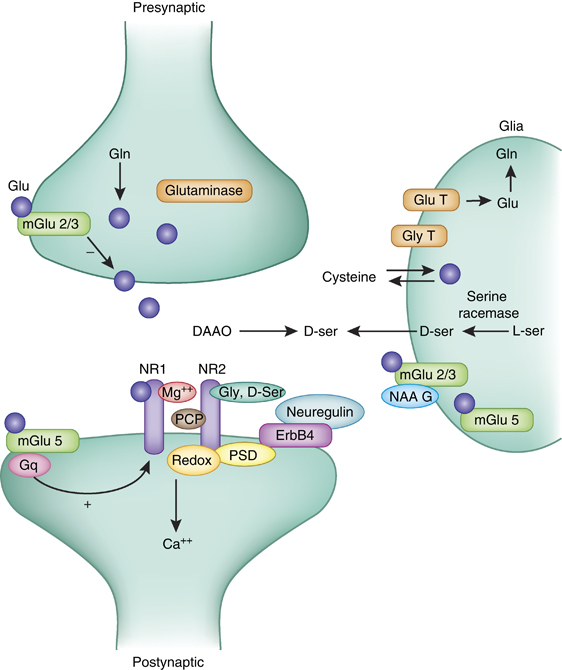
A simplified model of glutamate (Glu) synapse depicting some of the potential targets for manipulating the function of NMDA receptors. The two primary subunits of the receptor (NR1 and NR2) are depicted. On the presynaptic side, excess release of glutamate can be reduced by metabotropic group 2 receptors. Levels of vesicular glutamate also can be manipulated by the activity of the synthetic enzyme glutaminase, which converts glutamine (Gln) to Glu. On the postsynaptic site, several regulatory sites on the NMDA channel itself (eg, magnesium and PCP-binding sites, the D-serine and glycine (Gly) site and the redox (glutathione)) regulate the function of the receptor. In addition, other membrane-spanning receptors, such as the metabotropic group 5 (mGlu5) receptor or the ErbB4 receptor, indirectly influence the function of NMDAR by interacting through postsynaptic density (PSD) or signal transduction mechanisms. The glia includes a large number of proteins that influence both presynaptic and postsynaptic function of this synapse. These include transporters for both Glu and Gly, the D-serine-synthesizing enzyme serine racemase, D-serine transporter, as well as cystine–Glu transporter. In addition, a number of metabotropic Glu receptors including mGluR3 and mGluR5 are expressed by glia.
Dopamine Hypothesis of Schizophrenia: Version III
Presynaptic Glu hypothesis
Postsynaptic Glu hypothesis
Want to create your own Notes for free with GoConqr? Learn more.