Description
|
|
Created by Cher Bachar
over 12 years ago
|
|
Page 1
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
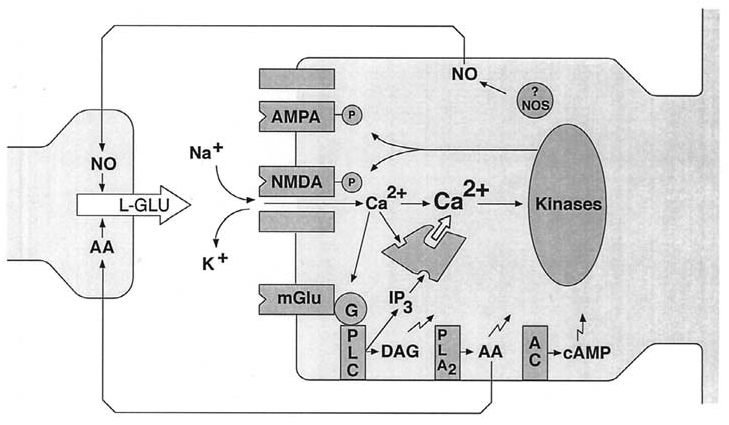
Figure 10–8. Signal transduction mechanisms in early LTP. The trigger for the induction of LTP is Ca2 permeating through the NMDA receptor. This is amplified by Ca2 release from Ca2/IP3- sensitive stores. The amplified Ca2 signal together with other activators of protein kinases (zigzag arrows) then leads to the phosphorylation of substrate proteins, including AMPA and NMDA receptors, and the insertion of AMPA receptors into the postsynaptic membrane (not shown). These post-translational processes contribute to E-LTP. Protein synthesis-dependent mechanisms at the synapse and by signals passing to the nucleus are responsible for L-LTP. Putative retrograde messengers, Nitric oxide (NO) and arachidonic acid (AA), that may stimulate presynaptic changes are also indicated. (Source: After Bliss and Collingridge, 1993.)
{kind=link}
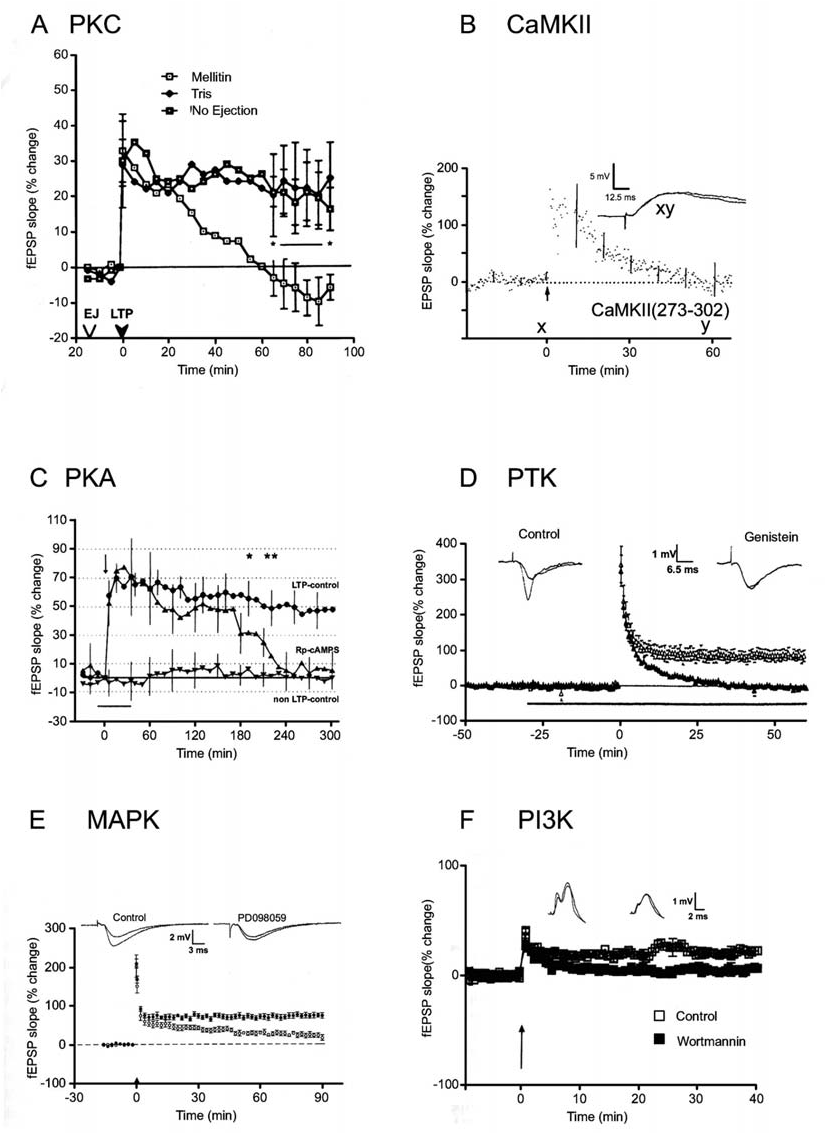
Figure 10–9. Role of protein kinases in the induction and expression of LTP. A. The PKC inhibitor mellitin, injected into the dentate gyrus in vivo 15 minutes prior to a tetanus, blocks E-LTP but spares STP. Similar findings were obtained with polymyxin B and H-7, implicating PKC as the kinase responsible. (Source: Lovinger et al., 1987.) In this and subsequent panels the slope of the fEPSP (or intracellular EPSP in B) is plotted versus time. Control experiments are also shown, except in B. In each case, a tetanus was delivered at t 0. B. A peptide inhibitor of CaMKII [CaMKII(273-302)] loaded into CA1 neurons in vitro blocks E-LTP but not STP. Similar results were obtained using a peptide inhibitor of PKC. (Source: Malinow et al., 1989.) C. The PKA inhibitor Rp-cAMPS blocks L-LTP in the CA1 region of the hippocampus in vitro. STP and E-LTP are unaffected. (Source: Matthies and Reymann, 1993.) D. The tyrosine kinase inhibitor genistein blocks E-LTP but not STP at CA1 synapses in vitro. Similar results were obtained using lavendustin A. (Source: O’Dell et al., 1991a.) E. The MAPK inhibitor PD098059 blocks induction of E-LTP but not STP at CA1 synapses in vitro. (Source: English and Sweatt, 1997.) F. ICV application of the PI3K inhibitor wortmannin blocks induction of E-LTP at perforant path-granule cell synapses in vivo. (Source: Kelly et al., 2000.)
The principal message to emerge from studies investigating the role of kinases in NMDA receptor-dependent LTP is that several kinases are involved but none is obligatory. The situation is complicated by the fact that the kinase cascades underlying LTP change during development. In area CA1 of the adult animal, as we have seen, CaMKII is required for the induction of LTP, and LTP can be induced in the presence of PKA inhibitors. Earlier, at postnatal day 9, the lead role is taken by PKA, as inhibitors of PKA, but not of CaMKII, block induction of LTP (Yasuda et al., 2003; see Fig. 10–21C). The situation is made even more complex by the existence of parallel kinase cascades, as first identified at CA1 synapses in juvenile rats. Thus at 2 weeks of age, a combination of inhibitors is required to block the induction of LTP.
Inhibitors of CaMKII, PKA, and PKC have no effect on induction when applied alone. Similarly, a PKA inhibitor plus a PKC inhibitor is ineffective. However, a CaMKII inhibitor applied together with either a PKA or a PKC inhibitor fully blocks the induction of LTP (Wikström et al., 2003). This suggests that two parallel kinase pathways, one involving CaMKII and the other involving both PKA and PKC, can each mediate LTP. As the animal matures, CaMKII assumes the dominant role, and PKA and PKC become more modulatory in nature. The involvement of parallel kinase cascades is, however, not simply a developmental issue, since a similar parallel pathway hasbeen observed in the medial perforant path in the dentate gyrus of adult mice (Cooke et al, 2006) and rats (Wu et al., 2006). Here we have a pathway that requires activation of CaMKII for one limb and PKA/MAPK for the other. Since both PKA and PKC can lead to activation of MAPK it is possible that in both juvenile CA1 and adult dentate gyrus the same two parallel pathways co-exist, one arm using CaMKII and the other PKA/PKC/MAPK.
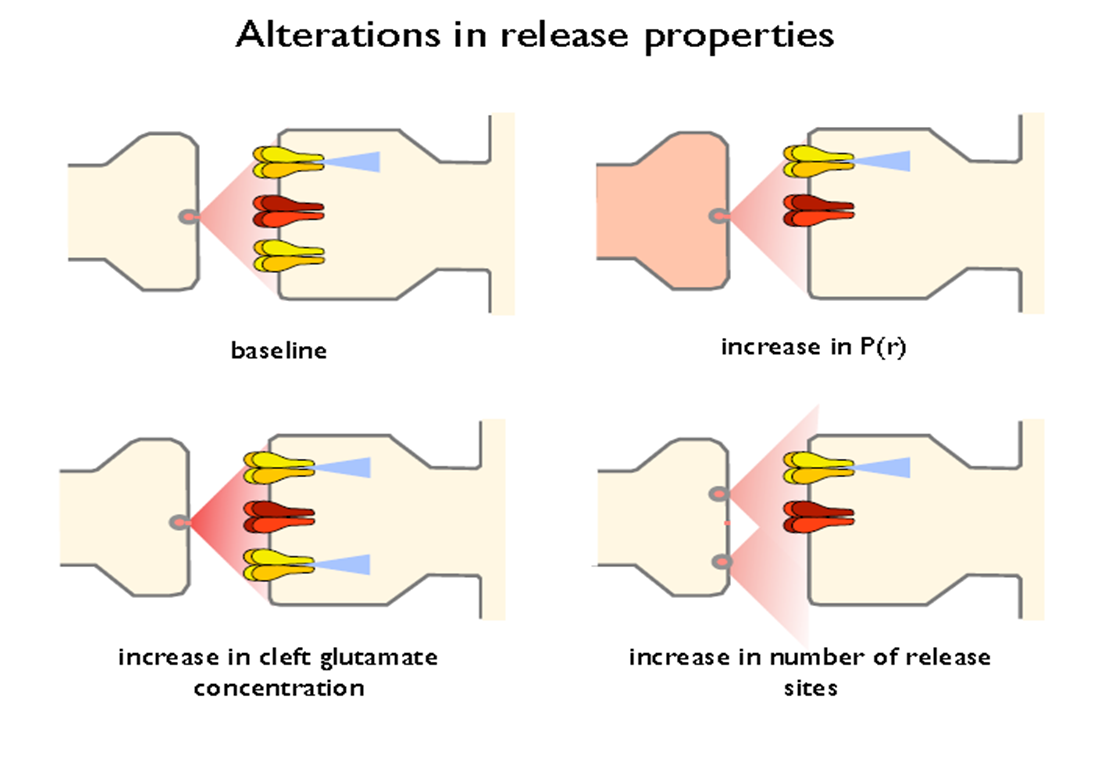
presynaptic- changes in release properties
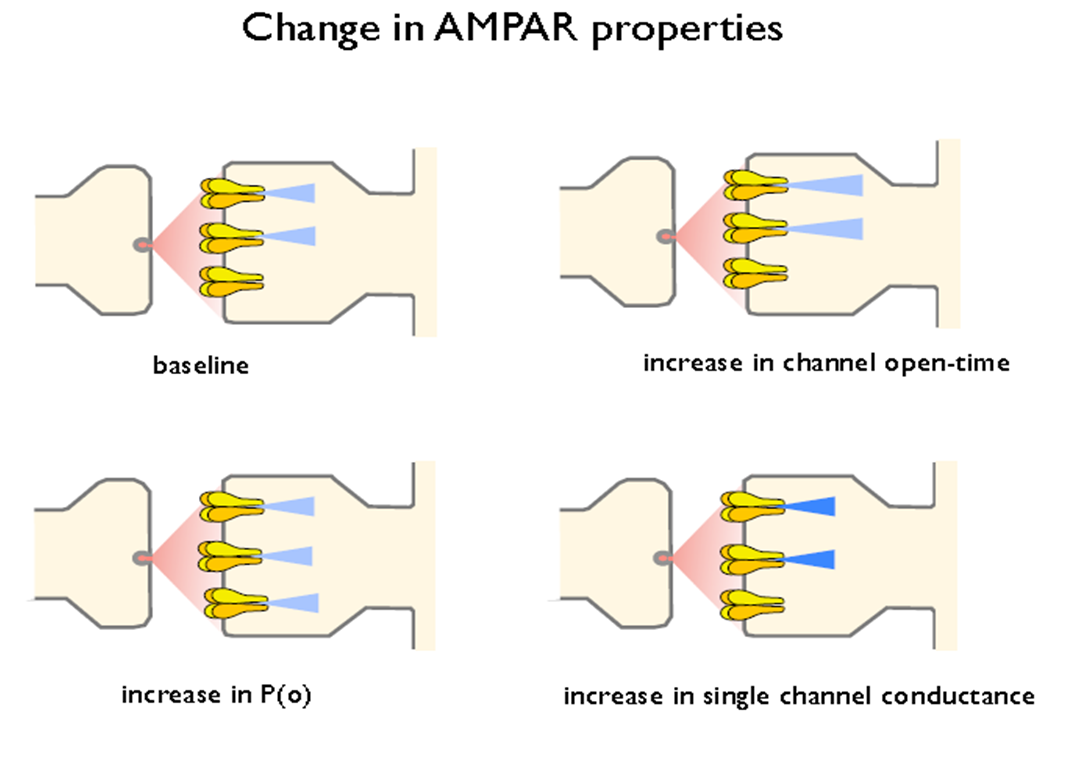
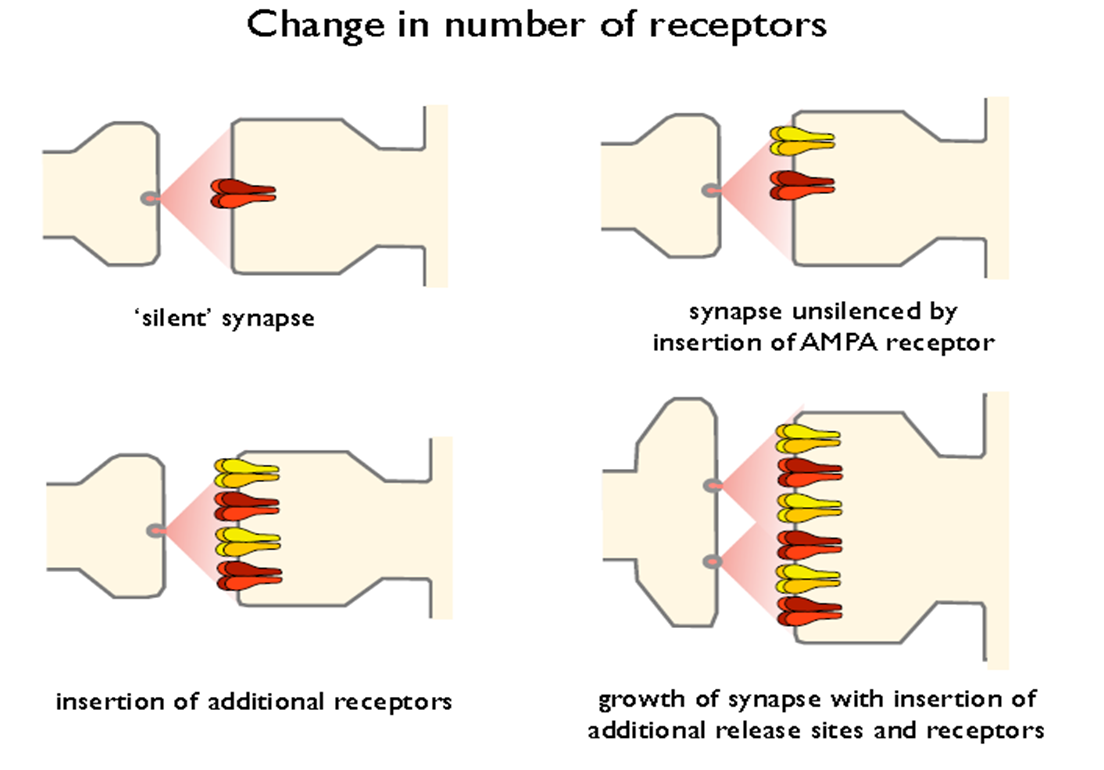
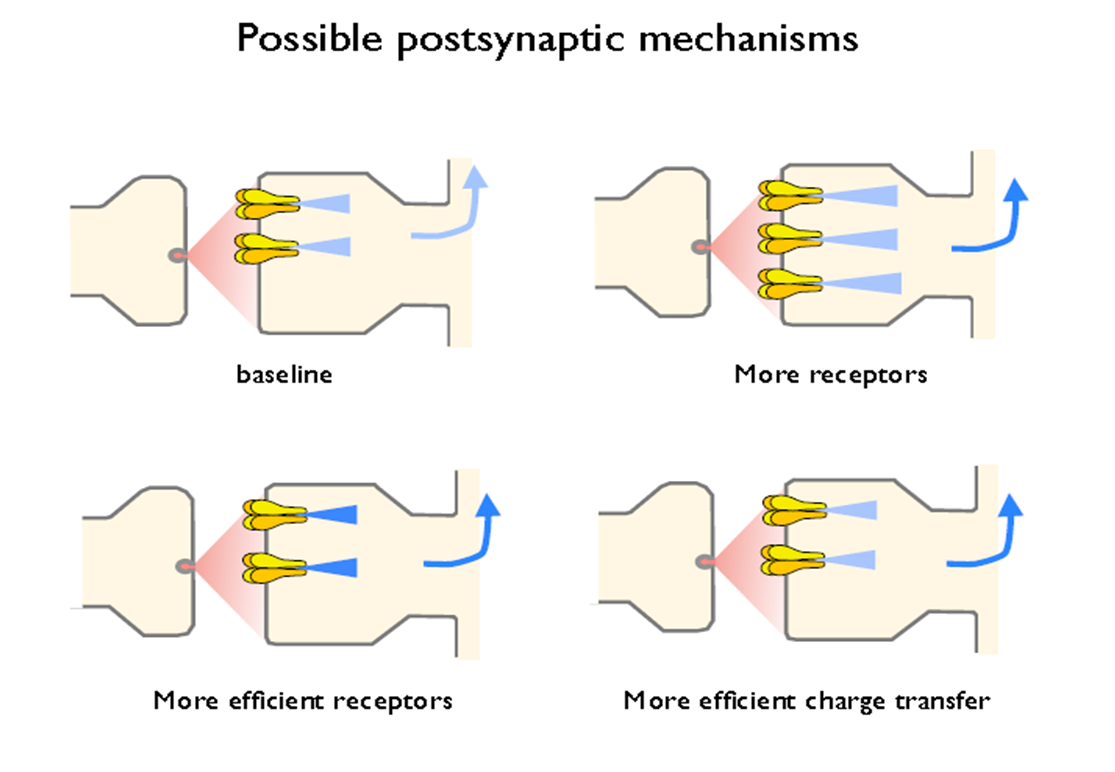
postsynaptic- no. of recep; AMPAR
early LTP signalling
data for protein kinases
multiple/parallel pathways- kinases
Want to create your own Notes for free with GoConqr? Learn more.