795954
Description
Flashcards by Heather Mansfield, updated more than 1 year ago
|
|
Created by Heather Mansfield
over 10 years ago
|
|
| Question | Answer |
| Things that need to be interchanged between an organism and the environment | Respiratory gases (oxygen and carbon dioxide). Nutrients (glucose, fatty acids, amino acids, vitamins, minerals). Excretory products (urea and carbon dioxide). Heat. |
| Surface area to volume ratio | Small organisms, have a surface area that is large enough, compared with their volume, to allow efficient exchange across their body surface. As organisms become larger their volume increases at a faster rate than their surface area. To overcome this organisms have evolved to either have a flattened shape so that no cell is ever far from the surface, or specialised exchange surfaces with large areas to increase the surface area to volume ratio. |
| Features of specialised exchange surfaces | - Large surface area to volume ratio to increase rate of exchange. - very thin so that the diffusion distance is short and therefore materials cross the exchange surface rapidly. - partially permeable to allow selected materials to cross without obstruction. - movement of the environmental medium eg. air, to maintain a diffusion gradient. - movement of the internal medium eg. blood, to maintain a diffusion gradient. - as they are thin they are easily damaged and therefore often located inside organisms, a method of moving the external medium over the surface eg. ventilation, is therefore needed. |
| Gas exchange in single-celled organisms | Single-celled organisms are small and therefore have large surface area to volume ratios. Oxygen is absorbed by diffusion across their body surface which is covered only by a cell-surface membrane. Carbon dioxide diffuses out across the surface in the same way. If the cell is surrounded by a cell wall, this is completely permeable and therefore not a barrier to gas diffusion. |
| Features of terrestrial insects to reduce water loss | Waterproof covering over their body surfaces - a rigid outer skeleton covered with a waterproof cuticle. Small surface area to volume ratio to minimize the area over which water is lost. |
| Gas-exchange system in insects | There is an internal network of tubes called tracheae. The trachea are supported by strengthened rings to prevent them from collapsing. The trachea divide into smaller tubes called tracheoles. The tracheoles extend throughout all the body tissues of the insect. |
| Use of diffusion in insects | When cells are respiring, oxygen is used up so its concentration towards the ends of the tracheoles falls. This creates a diffusion gradient that causes oxygen to diffuse from the atmosphere along the trachea and tracheloes to the cells. Carbon dioxide is produced during respiration. This creates a diffusion gradient in the opposite direction. Carbon dioxide diffuses along the tracheoles and trachea from the cells to the atmosphere |
| Use of ventilation in insects | The movement of muscles in insects can create mass movements of air into and out of the trachea. This further speeds up the exchange of respiratory gases. |
| Spiracles in insects | Spiracles are tiny pores on the body surface. The spiracles may be opened and closed by a valve. When the spiracles are open, water can evaporate from the insect. For much of the time insects keep their spiracles closed to prevent this water loss. Periodically the spiracles open to allow gas exchange. |
| Limitations of the insect gas exchange system | It relies mostly on diffusion to exchange gases between the environment and the cells. For diffusion to be effective, the diffusion pathway needs to be short. As a result this limits the size that insects can attain. |
|
Image:
Picture1 (image/png)
|
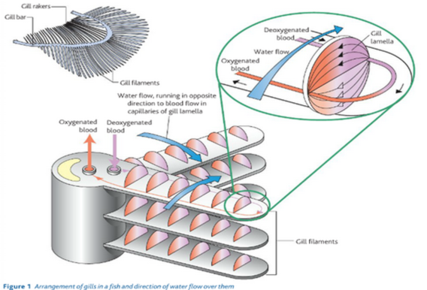
Structure of the gills: The gills are located in the body of the fish behind the head. They are made up of gill filaments. The fill filaments are stacked up in a pile. At right angles to the gill filament are fill lamellae, which increase the surface area of the gills. Water is taken in through the mouth and forced over the gills and out through an opening on each side of the body. |
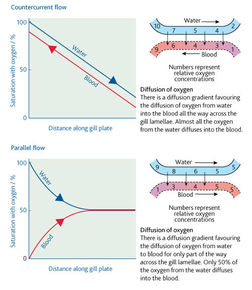
| Countercurrent flow | The flow of water in the gill lamellae and the flow of blood within them are in opposite directions. This is important for ensuring that the maximum possible gas exchange is achieved. If the water and blood flow in the same direction, far less gas exchange would take place. |
| The arrangement of a counter current system means that blood that is already loaded with oxygen meets water, which has its maximum concentration of oxygen. Therefore diffusion of oxygen from the water to the blood takes place. Blood with little or no oxygen in it meets water which has has most but not all, of its oxygen removed. Again diffusion of oxygen from the water to the blood takes place. There is therefore a fairly constant rate of diffusion across the entire length of the gill lamellae. In this way, about 80 per cent of the oxygen available in the water is absorbed into the blood of the fish. | |
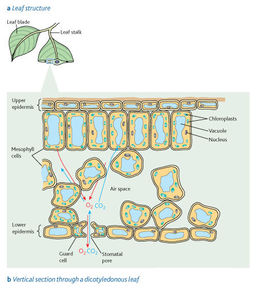
| Gas exchange in the leaf of a plant | Like animal cells, all plant cells take in oxygen and produce carbon dioxide during respiration. However some plant cells carry out photosynthesis, during this process the plant cell takes in carbon dioxide and produces oxygen. At times the gases produced in one process can be used for the other which reduces the need for gas exchange with the external air. When photosynthesis is taking place all though some carbon dioxide comes from respiration most of it has to be obtained from the external air and most of the oxygen produced diffuses out of the plant, although some is used for respiration. When photosynthesis is not occurring oxygen diffuses into the leaf and carbon dioxide diffuses out. |
| Structure of a plant leaf | |
| Adaptation of a leaf for gas exchange | No living cell is far from the external air, and therefore a source of oxygen and carbon dioxide. Diffusion takes place in the gas phase (air), which makes it more rapid than if it were in water. There is a short, fast diffusion pathway. The plant leaf has a very large surface area compared with the volume of living tissue. Diffusion gradients are maintained by the mitochondria carrying out respiration and chloroplasts carrying out photosynthesis. |
| Stomata | Stomata are minute pores, which occur mainly on the leaves, especially the underside. Each stoma is surrounded by a pair of guard cells which can open and close the stomatal pore. In this way they can control the rate of gaseous exchange. This is important because plants lose water by evaporation, plants have to balance this with the conflicting need for gas exchange. Stomata are completely or partly closed at times when water loss would be excessive. |
| Why do large organisms need a transport system? | Exchange surfaces are located in specific regions of the organism. A transport system is required to take materials from cells to exchange surfaces and visa versa. Whether or not there is a specialised transport medium and whether it is circulated by a pump depends on the surface area to volume ratio and how active the organism is. |
| Features of transport systems | * A suitable medium in which to carry materials, this is normally a liquid based on water because water readily dissolves substances and can be moved around easily * A form of mass transport in which the transport system is moved around in bulk over large distances * A closed system of tubular vessels that contains the transport medium and forms a branching network to distribute it to all parts of the organism * a mechanism for moving the transport medium within vessels, this requires a pressure difference which is achieved through contraction of muscles (animals) or water evaporation (plants * a mechanism to maintain the mass flow movement in one direction * a means of controlling the flow of the transport medium to suit the changing needs of different parts of the organism. |
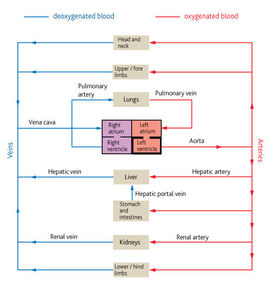
| Transport system in mammals | Blood is confined to vessels. A muscular pump called the heart circulates blood around the body. Mammals have a double circulatory system to maintain pressure. Substances are delivered to the rest of the body quickly which is necessary due to the high rate of metabolism in mammals. The vessels of mammals are divided into arteries, veins and capillaries. Although the transport system moves substances long distances the final part of the journey is by diffusion. |
| Mammalian circulatory system | |
| Types of blood vessles | Arteries carry blood away from the heart and into arterioles. Arterioles are smaller arteries that control blood flow from the arteries to the capillaries. Capillaries are tiny vessels that link arterioles to veins. Veins carry blood from capillaries back to the heart. |
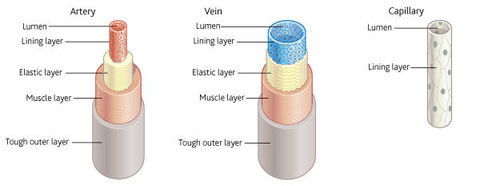
| Basic structure of blood vessles | Tough outer layer that resists pressure changes from both within and outside. Muscle layer than can contract and so control the flow of blood. Elastic layer that helps to maintain blood pressure by stretching and springing back. Thin inner lining (endothelium) that is smooth to prevent friction and thin to allow diffusion. Lumen - the central cavity of the blood vessel through which the blood flows |
| Structure of arteries, veins and capillaries | |
| Artery structure related to function | The function of arteries is to transport blood rapidly under high pressure from the heart to the tissues. The muscle layer is thick compared to veins. This means smaller arteries can be constricted and dilated in order to control the volume of blood passing through them. The elastic layer is relatively thick compared to veins because it is important that the blood pressure in the arteries is kept high if blood is to reach the extremities of the body. The elastic wall is stretched at each systole and spring back during diastole. The stretching and recoil helps maintain a high pressure. The overall thickness of the wall is large to resist the vessel bursting under pressure. There are no valves because the blood is under constant high pressure and therefore does not tend to flow backwards |
| Arteriole structure related to function | Arterioles carry blood under lower pressure than arteries, from arteries to capillaries. They also control the flow of blood between the two. The muscle layer is relatively thicker than in arteries, the contraction of the muscle layer allows constriction of the lumen of the arteriole, which restricts the flow of blood and so controls its movement into the capillaries that supply the tissues with blood. The elastic layer is relatively thinner than in arteries because blood pressure is lower. |
| Vein structure related to function | Veins transport blood slowly, under low pressure, from the tissues to the heart. The muscle layer is relativley thin compared to arteries because veins carry blood away from the tissues and therefore their constriction and dilation cannot control the flow of blood to the tissues. The elastic layer is relatively thin compared to arteries because the low pressure of blood within the veins will not cause them to burst and pressure is too low to create a recoil action. The overall thickness of the wall is small because there is no need for a thick wall as pressure within the veins is too low to create any risk of bursting. It also allows them to be flattened easily, aiding the flow of blood within them. There are valves throughout to ensure that blood does not flow backwards, which it might otherwise do because the pressure is so low. When body muscles contract, veins are compressed, pressurising blood within them. The valves ensure that the pressure directs blood in one direction only. |
| Capillary structure related to function | The function of the capillaries is to exchange metabolic materials such as oxygen, carbon dioxide and glucose between the blood and the cells of the body. The flow of blood in the capillaries is much slower to allow more time for the exchange of materials. Their walls consist of only the lining layer making them extremely thin to distance over which diffusion takes place is short. This allows for rapid diffusion of materials between the blood and the cells. They are numerous and highly branches, thus providing a large surface area for diffusion. They have a narrow diameter and so permeate tissues, which means that no cell is far from a capillary. Their lumen is so narrow that red blood cells are squeezed against the side of a capillary. This brings them even closer to the cells to which they supply oxygen. This again reduces diffusion distance. There are spaces between the lining cells that allow white blood cells to escape in order to deal with infections within tissues. |
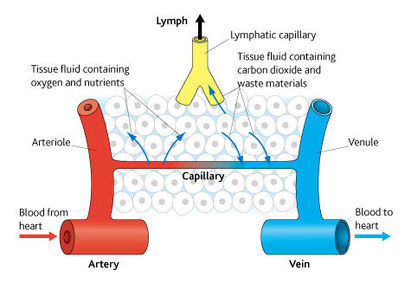
| What is tissue fluid? | Tissue fluid is a watery liquid that contains glucose, amino acids, fatty acids, salts and oxygen. Tissue fluid supplies all of these substances to the tissues. In return, it receives carbon dioxide and other waste materials from the tissues. Tissue fluid is therefore the means by which materials are exchanged between blood and cells, it bathes all the cells of the body. It is formed from blood plasma, which is controlled by homeostatic systems. As a result tissue fluid provides a constant environment for the cells it surrounds. |
| Formation and return of tissue fluid | |
| How does tissue fluid form | Blood is pumped by the heart into narrower and narrower vessels. This creates hydrostatic pressure, at the arterial ends of the capillaries. This hydrostatic pressure forces tissue fluid out of the blood plasma. The outward pressure is however opposed by the force of the hydrostatic pressure of the tissue fluid outside the capillaries and the lower water potential of the blood due to the plasma proteins which pulls water back into the blood within the capillaries. The combined effect of all these forces creates an overall pressure that pushes tissue fluid out of the capillaries. This pressure is only enough to force small molecules out of the capillaries, leaving all the cells and proteins in the blood - this is ultra-filtration. |
| Return of tissue fluid to the circulatory system via capillaries | Most tissue fluid returns to the blood plasma via the capillaries. The loss of tissue fluid from the capillaries reduces the hydrostatic pressure inside them. As a result by the time the blood has reached the venous end of the capillary network its hydrostatic pressure is less than that out the tissue fluid outside it. Therefore tissue fluid is forced back into the capillaries by the higher hydrostatic pressure outside them. In addition, the osmotic forces resulting from the proteins in the blood plasma pull water back into the capillaries. The tissue fluid has lost much of its oxygen and nutrients by diffusion into the cells that it bathed, but it has gained carbon dioxide and waste materials in return. |
| Return of tissue fluid to the circulatory system via lymphatic system | Not all the tissue fluid can return to the capillaries, the remainder is carried back via the lymphatic system. This is a system of vessels that begin in tissues. Initially they resemble capillaries, but they gradually merge into larger vessels that form a network throughout the body. These larger vessels drain their contents back into the bloodstream via two ducts that join veins close to the heart. The contents of the lymphatic system are moved by hydrostatic pressure of the tissue fluid that has left the capillaries and contraction of body muscles which squeeze the lymph vessels, valves in the lymph vessels ensure that the fluid inside them moves away from the tissues in the direction of the heart |
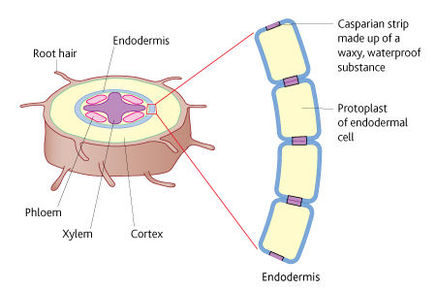
| Root of a dicotyledonous plant | |
| Uptake of water by root hairs | Each root hair is a long, thin extension of a root epidermal cell. They are efficient surfaces for exchange because they provide a large surface area as they are long and occur in thousands in the branches of a root and have a thin surface layer across which materials can move easily. The soil solution has a high water potential, the root hairs have a much lower water potential so water moves by osmosis from the soil solution into the root-hair cells down a water potential gradient. |
| The apoplastic pathway | As water is drawn into endodermal cells, it pulls more water along behind it due to the cohesion of water molecules. This creates a tension that draws water along the cell walls of the cells of the root cortex. The structure of the cell walls has many water filled spaces so there is little resistance to the pull of water along the cell walls. |
| The symplastic pathway | This taken place across the cytoplasm of the cells of the cortex as a result of osmosis. The water passes through the cell walls along tiny openings called plasmodesmata. Each plasmodesma is filled with a thin strand of cytoplasm. There is therefore a continuous column of cytoplasm extending from the root hair cell to the xylem at the center of the root. |
| Symplastic pathway steps | Water entering by osmosis increases the water potential of the root-hair cell. The root-hair cell now has a higher water potential than the first cell in the cortex. Water therefore moves from the root-hair cell to the first cell in the cortex by osmosis, down the water potential gradient. The first cell now has a higher water potential that its neighbour to the inside of the stem. Water therefore moves into this neighbouring cell by osmosis along the water potential gradient. The second cell now has a higher water potential than its neighbor to the inside, and so water moves from the second cell to the thirds cell by osmosis along the water potential gradient. At the same time, this loss of water from the first cortical cell lowers its water potential, causing more water to enter it by osmosis from the root-hair cell. In this way, a water potential gradient is set up across the cells of the cortex, which carries water long the cytoplasm from the root-hair cell to the endodermis. |
| The casparian strip | When water reaches the endodermis by the apoplastic pathway, the waterproof band that makes up the Casparian strip in endodermal cells prevents it progressing further along the cell wall. At this point, water is forced into the living protoplast of the cell, where it joins water that has arrived there by the symplastic pathway. |
| Active transport | Active transport of salts is the most likely mechanism by which water now gets into the xylem. Endodermal cells actively transport salts into the xylem, this process can only take place within living tissue because energy is required. It takes place along carrier proteins in the cell-surface membrane. If water is to enter the xylem, it must first enter the cytoplasm of the endodermal cells. The active transport of mineral ions into the xylem by the endodermal cells creates a lower water potential in the xylem, by osmosis, along a water potential gradient. This water potential gradient is the result of the active transport of salts into the xylem from the endodermal cells. This creates a force that helps to move water up the plant, this force is called root pressure. |
| Evidence for root pressure | * The pressure increases with a rise in temperature and decreases at lower temperatures. * Metabolic inhibitors, such as cyanide, prevent most energy release by respiration and also cause root pressure to cease. * A decrease in the availability of oxygen or respiratory substrates causes a reduction in root pressure. |
| Movement of water out through stomata | The humidity of the atmosphere is usually less than that of the air spaces next to the stomata. Provided the stomata are open, water molecules diffuse out of the air spaces into the surrounding air. Water lost from the air spaces is replaced by water evaporating from the cell walls of the surrounding mesophyll cells. By changing the size of the stomatal pores, plants can control their rates of transpiration |
| Movement of water across the cells of a leaf | Water is lost from the mesophyll cells by evaporation from their surfaces to the air spaces of the leaf. This is replaces by waer reaching the mesophyll cells from the xylem by either the apoplastic or symplastic pathways. In the case of the symplastic pathway the water movement occurs because the mesophyll cells lose water to the air spaces, these cells now have a lower water potential and so water enters by osmosis from neighbouring cells, the loss of water from these neighbouring cells lowers their water potential, they, in turn, take in water from their neighbours by osmosis. In this way, a water potential gradient is established that pulls water from the xylem, across the leaf mesophyll, and finally out into the atmosphere. |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Want to create your own Flashcards for free with GoConqr? Learn more.