17778967
4. Membrane Structure and Function
Description
No tags specified
Slide Set by Chloe Cavarretta, updated more than 1 year ago
More
Less
|
|
Created by Chloe Cavarretta
almost 7 years ago
|
|
Resource summary
Slide 1
Learning Outcomes
Describe the structural features of different lipids and appreciate how these features enable the formation and variability of biological membranes
Understand the significance of the fluid mosaic model of biological membranes – how it explains the features necessary for a functioning biological membrane and what its limitations are
Evaluate the molecular features and energetic barriers for lipid transport between and across membranes
Appreciate how proteins interact with biological membranes
Understand how different classes of molecules cross biological membranes
Slide 2
Lipid components
Lipids spontaneously assemble into different structures in aqueous environment
Driving force is exclusion of water from hydrophobic chains
Structures are:
Bilayer
Micelles
Liposomes
Slide 3
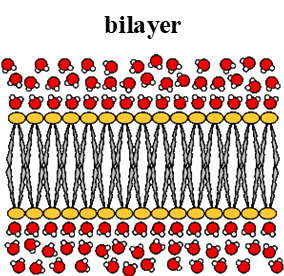
1. Bilayer
Biological membranes are made of lipid bilayer of amphipathic PLs: PLs have charged head group and 2 hydrophobic tails
Non-amphipathic lipids include fats: triacylglyceride and triacylglycerol
Shape= sheet-like
{kind=link}
Slide 4
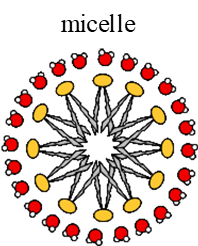
2. Micelles
Monolayer (not bilayer- do not make membranes)
Inner hydrophobic environment
{kind=link}
Slide 5
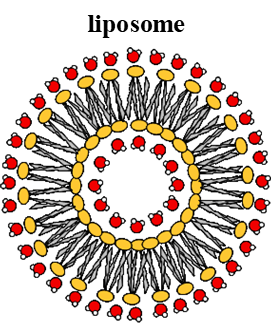
3. Liposomes
{kind=link}
Form bilayer
Inside and outside is hydrophillic aqueous environment
Slide 6
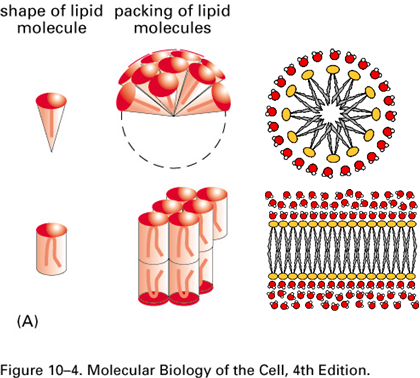
Lipid shape and membrane curvature
Relative cross-sectional area of head:tails determine lipid shape and lipid shape determines curvature
Conical Shape:
Larger head, smaller tail CS-area
Create curved membrane
Cylindrical Shape:
Head and tail occupy similar area
Create sheet shaped membrane
{kind=link}
Slide 7
Sheet formation vs liposome
Both bilayers
End of the sheet is highly unfavourable because tails are exposed to water: sheet folds on itself to bury tails
Liposome forms when:
Closure happens quickly from start of formation of sheet, sheet isnt too big so curved ends meet
Plasma membrane forms when:
Closure happens far away from beginning of sheet formation
Large, extended sheet like structure
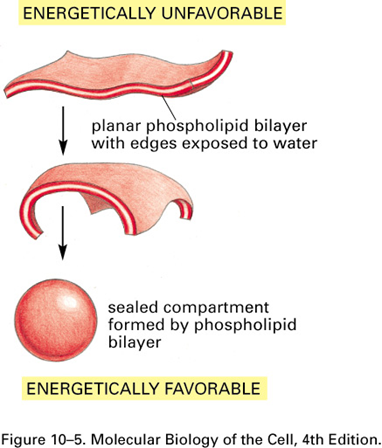{kind=link}
Slide 8
Fluidity and membrane thickness
Bilayers either have a gel-like or fluid-like consistency
Gel-like consistency (thicker):
Lower temperatures
Lipids in each half of bilayer arranged in 2D lattice
More ordered and tightly packed
Mainly saturated lipids
Fluid-like consistency (thinner):
At temperatures we live at
Liquid crystalline state
Less ordered and less tightly packed
Mainly unsaturated lipids
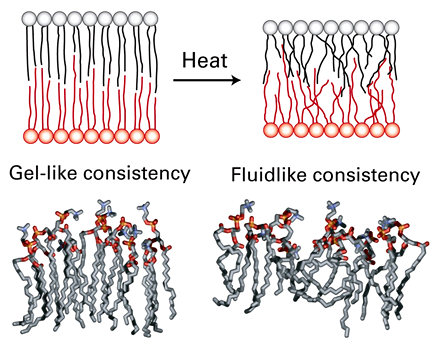{kind=link}
Slide 9
Fluidity and membrane thickness
Composition of saturated and unsaturated lipids determine the fluidity:
Unsaturated lipids disrupt the ordered packing of saturated lipids
Bilayer has more free space and is more permeable to water
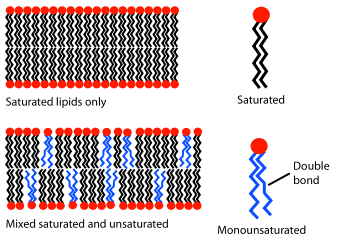{kind=link}
Slide 10
Fluidity and membrane thickness
Sterols (eg cholesterol) affect membrane by:
Fluidity decreases, increase packing and make more rigid
Permeability to neutral solutes and ions decreases
Thickness increases
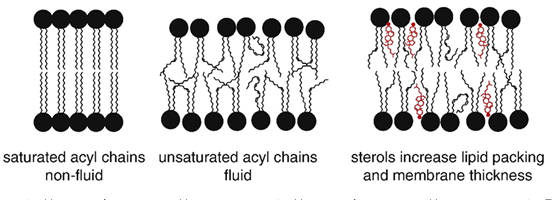{kind=link}
Slide 11
Fluid mosaic model
Lipid bilayer gives the fluidity and there are proteins embedded within bilayer
Model allows:
Lipid movement in plane of bilayer
Membrane plasticity/deformability (easily shaped)
Protein functions (transport, enzyme activity, conformational change)
Slide 12
Fluid mosaic model: Old experiment
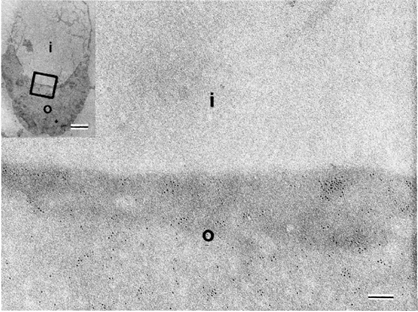{kind=link}
Caption: : Electron micrograoh of rabbit erythrocyrte membrane stained with ricin Singer and Nicoloson 1972
Erytrocytes stained with ricin
Ricin binds to sugars on surface of PM (darker, more e dense)
PM in is folded, both inside and outside surfaces are visible in same image, can see differences in ricin binding
Proves that 2 sides of membrane are different so membrane is asymmetrical and shows clustering of membrane components
Slide 13
Fluid mosaic model: Recent experiment
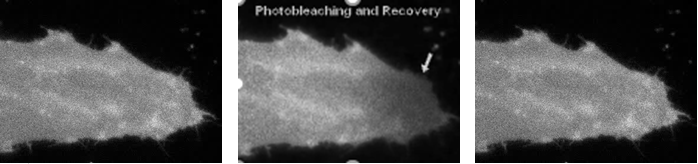{kind=link}
Label cell with fluorophore
Shine laser light onto one part of cell membrane, bleaches it and destroys its fluorescence
Fluorescence only recovers if there is diffusion into the region that was bleached
Diffusion of proteins within the membrane is evidence for fluidity
Slide 14
Different shapes of biological membranes
ER:
Made of sheets and a network of tubules
Sheets and tubules use different proteins/ lipids to shape them differently- eg reticulon proteins curve membrane to shape tubules, also found at edges of ER sheets to tightly curve
Different functions: Sheets have ribosomes = protein synthesis, tubules(smooth ER)= lipid synthesis and secretion
Golgi:
Made of stack of cisternae (like sheet) and vesicles (like tubules)
Vesicles= smaller tightly curved structures than sheets, curved in 3D(tubules curved in 2D)
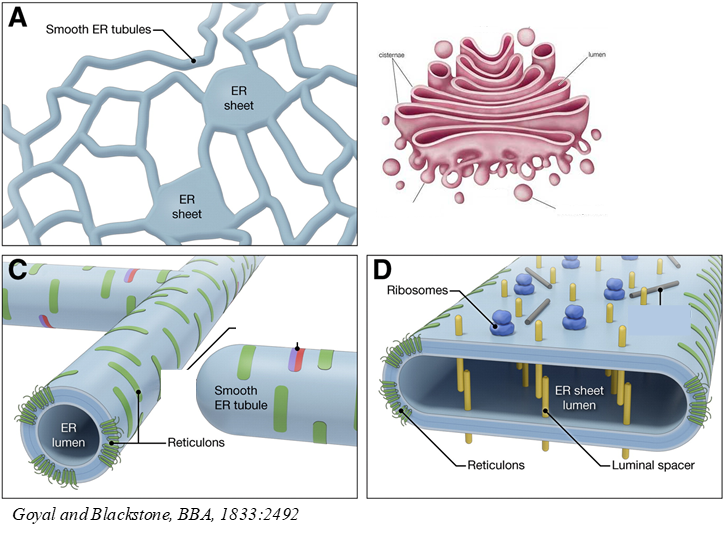{kind=link}
Slide 15
Membrane deformation: protein help
Amphipathic helices:
Hydrophillic and hydrophobic aa on opposite sides of helix (phillic facing aq, phobic in membrane)
Wedges into bilayer between heads, curving bilayer
Loop insertion:
Loops with hydrophobic amino acids that form a larger wedge in membrane, more curvature
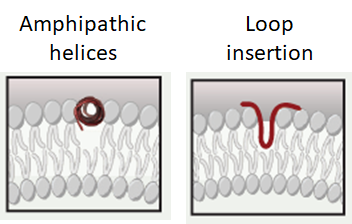{kind=link}
Slide 16
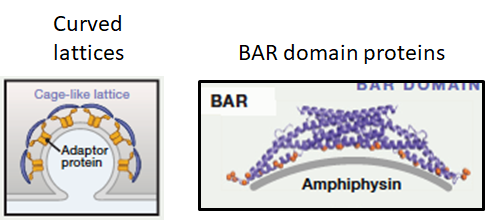
Curved lattice:
Lattices bind cargo proteins whilst forming curved polymer
Protein complex to shape membrane, vary in curvature
BAR domain proteins:
Proteins bind the bilayer via a curved surface eg BAR domain containing proteins
BAR domain- curved protein structure that binds curved side via lipid heads
{kind=link}
Slide 17
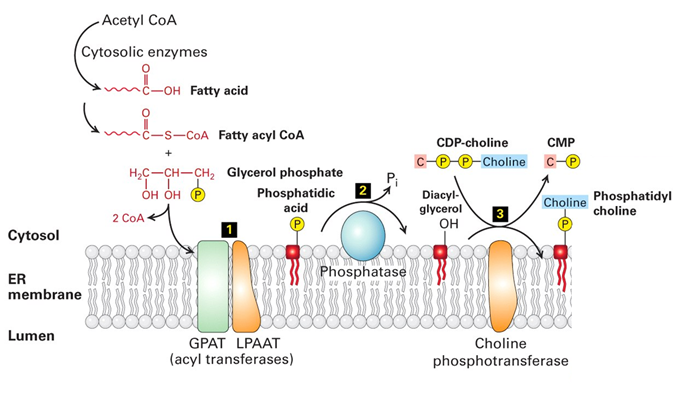
Transformation of lipids
Lipids are altered throughout the cell to fit the function of each membrane type
Lipid biosythesis is initiated in the smooth ER:
Fatty acid is synthesised in cytosol
ER membrane bound acyl transferases turn it into phosphatidic acid intermediate
Other membrane bound enzymes, transeferases put head groups on (eg choline to make phosphatidyl choline)
Other PLs (eg phosphatidyl serine) are made by a series of conversions from each other
{kind=link}
Slide 18
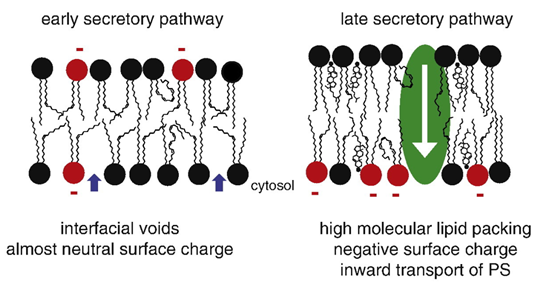
Lipid remodelling in the secretory pathway
Membranes have almost neutral surface charge and are almost fluid in early secretory pathway:
Unsaturated FA chains are replaced by more saturated FA from ER to PM
Glycolipids head groups synthesised into elaborate glycans in Golgi
Membranes become more asymmetrical towards PM
More cholesterol and sphingolipids in late pathway
Membrane thickness increases towards PM
Phosphatidyl serine (PS) flips onto the inside membrane
{kind=link}
Slide 19
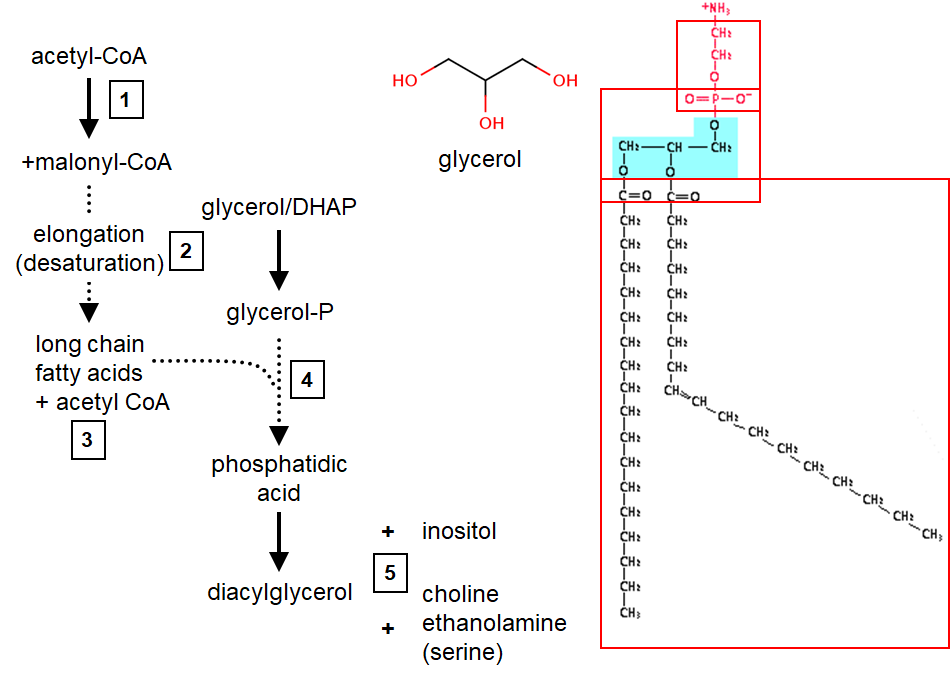
PL synthesis
{kind=link}
3 moieties make up a PL: glycerol, FA tail and head group
Overall process: (1) make FA, (2) modify FA, (3) stick 2nd FA on, (4) stick head group on eg inositol, serine, choline
FA synthesis in cytosol, involves the polymerisation of 2C building blocks
Enzymes: acetyl-CoA carboxylase(ACC)= carboxylates acteyl CoA(C2) malonylCoA(C3), FA synthase= uses 2 C of each malonylCoA to elongate chain (decarboxylative condensation reaction)
Slide 20
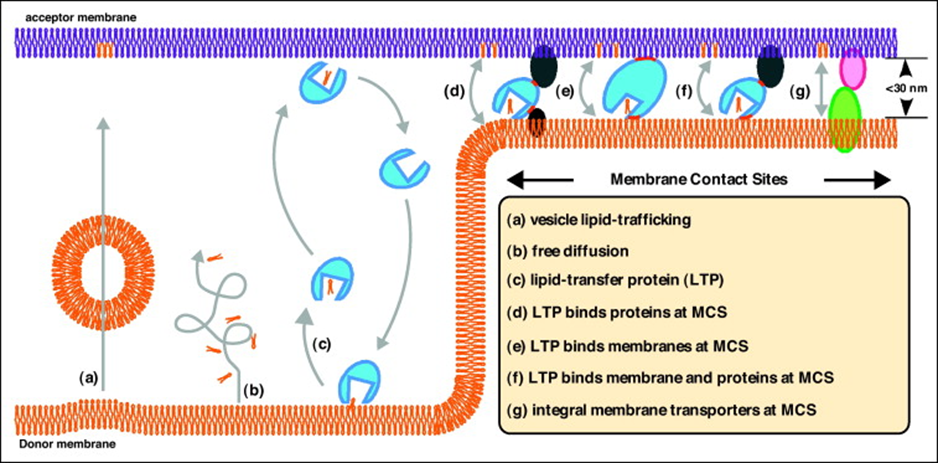
Transfer of PL between organelles
Lipids are transferred from the ER to target in 3 different ways(a-c):
Using vesicles
Free diffusion in cytosol (unfavourable due to hydrophobicity)
Lipid transfer proteins
Other means of transport(d-g):
Membrane contact sites (MCS)
{kind=link}
Slide 21
Membrane contact sites
When one organelle is in close proximity (<30nm) with another, intense lipid transfer by lipid transfer proteins that are:
In complex with other LTPs(d)
Bridging the membrane, binds membrane at both ends(e)
Integral membrane transporters that are integral to both membranes at MCS (g)
Other functions of MCS:
Lipid transfer
Coordinate Ca3+ release to facilitate signalling/cytoskeleton dynamics (between ER and PM)
Aid in organelle fission (between mitochondria and ER)
Aid in protein sorting within/between organelles
Slide 22
Lipid transfer proteins
Most LTPs can bind lipid monomers in a hydrophobic pocket to transfer hydrophobic lipids through an aqueous phase, eg CERT
LTPs show specificity for lipid types:
PC-TP (phosphatidyl choline transfer protein)
CERT (ceramide transfer protein)- moves ceramides with C14-C20
PL-TP (phospholipid transfer protein)- least selective, also present in blood plasma associated with HDL
{kind=link}
Slide 23
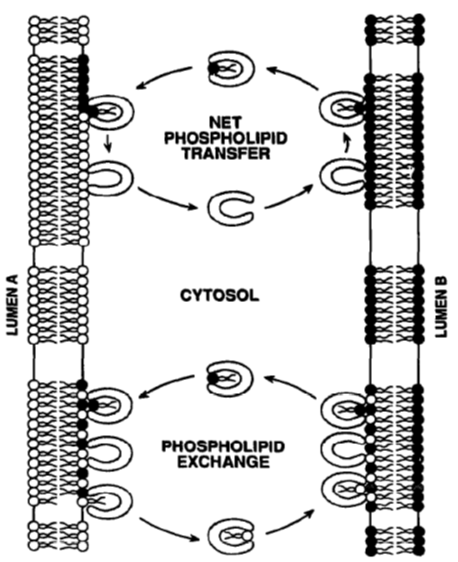
Lipid transfer proteins
LTPs work in 2 ways:
Net PL transfer
Transfer PL from one membrane to another, LTP goes back freely
Eg transfer from ER due to PL synthesis
PL exchange
Transfer PL in one direction and another in opposite direction
{kind=link}
Slide 24
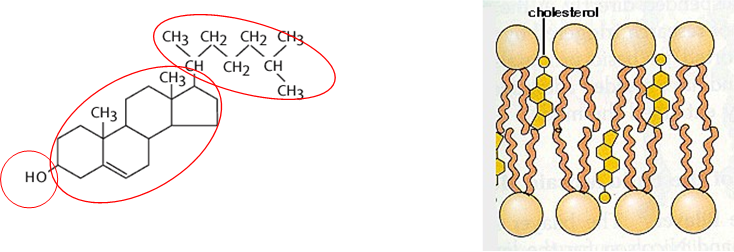
Cholesterol
Made of 3 components: Alcohol (OH interacts with heads of PL and sphingolipids), 4 ring steroid domain and lipid tail (both embedded in membrane, increase packing)
Chemical analogues in other organisms- ergosterol in yeast
Integral part of membrane rafts
How its made: AcetlyCoA in cytoplasm -> mevalonate in ER (C6) -> decarboxylation to isoprene (C5) -> 6X isoprene = cholesterol (C27) (subunit is isoprene)
{kind=link}
Slide 25
Cholesterol transport
Synthesis/uptake controlled by SREBPs (sterol regulatory element binding proteins): low levels of sterols = SREBP cleaved at Golgi to soluble form that translocates to nucleus and increases expression of genes for synthesis/uptake
Cholesterol supplied to body by: eating cholesterol containing foods (intestine) or biosynthesis (liver)
Long distance transport of cholesterol in blood plasma (bound to lipoproteins HDL and LDL):
Distributed from liver/intestine to tissues by LDL
LDL binds cholesterol, binds LDL receptors in tissues, endocytosed, hydrolysis in lysosome and released into tissues
LDL too high= too much cholesterol in blood, HDL transports in opposite direction (tissues to liver) for destruction
Balance between HDL and LDL
Slide 26
Lipid droplets
Predominantly found in adipose tissue
Storage of lipids: free lipids can be toxic to cell in large quantities (ROS)
Surrounded by PL monolayer, surface has proteins (loops/amphipathic helices) for formation, maintenance and regulation (NEVER TM, not happy with hydrophobicity)
Generated at the ER, site of lipid biosynthesis
Slide 27
Membrane proteins + glycolipids
Membranes also function as a structure for proteins:
Eg synaptic vesicle: dominated by proteins (200+ types), majority span the membrane,(vesicle lumen has high protein content) function in cell-cell communication, signalling, transport
Membrane fuzz= glycans (glycolipid is a type of glycan)
Glycolipid:
Made in ER and Golgi by stepwise addition of monosaccharides
First addition done on cytosolic/ lumenal faces of ER or Golgi
Glycolipid synthesis starts in ER with ceramide, elaboration with additional and branched oligosaccharides occurs in Golgi
Blood antigens are an example of glycolipids
Slide 28
Glycolipids
Gangliosides (glycosphingosides)= type of glycolipid important for brain function:
Highly elaborated
Essential component of neural membranes
Important component of lipid rafts
Face outside the cell
Used by pathogens for attachment/ entry into cell eg cholera toxin binds GM1 ganglioside
Slide 29
Summary
Describe the structural features of different lipids and appreciate how these features enable the formation and variability of biological membranes
Lipids can be cone or cylindrical shaped dependent on the head-group and tail. Lipid shape determines membrane shape – sheet vs tubule vs vesicle. Most lipid synthesis happens at the ER. Glycolipids are signature features of a cell’s exterior.
Understand the significance of the fluid mosaic model of biological membranes – how it explains the features necessary for a functioning biological membrane and what its limitations are
Allows diffusion in the membrane plane, permits asymmetry across bilayers. Membrane contact sites restrict membrane fluidity, creating novel biological functions.
Evaluate the molecular features and energetic barriers for lipid transport between and across membranes
Carrier proteins and short range protein shuttles can help lipids cross the cytosolic space between membranes. Membrane contact sites permit lipid transfer. Lipid droplets act as storage places for potentially toxic lipid
Slide 30
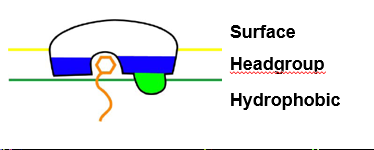
1. Peripheral membrane proteins
Non-permanent way to bind proteins to membrane (non-integral)
Proteins bind heads of PLs- anchors protein to membrane
Major structural features:
Polar pocket/ groove that recognises specific ligand
Hydrophobic protrusion that penetrates into membrane
Cluster of basic residues that bind anionic PL headgroup
{kind=link}
Slide 31
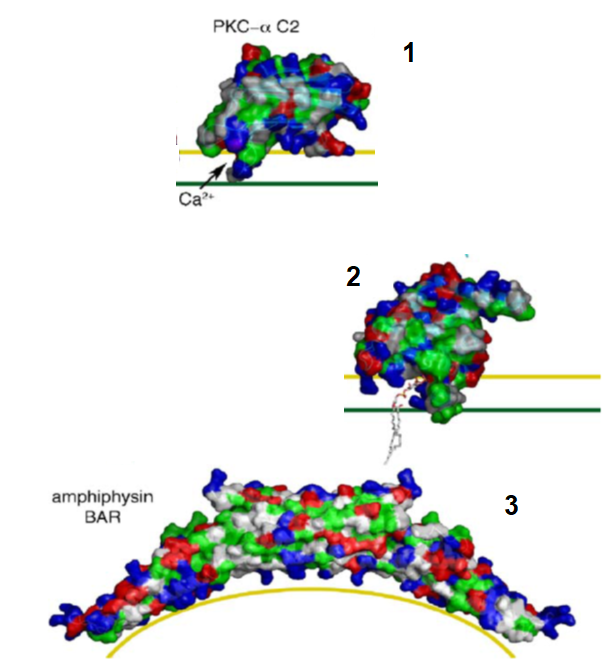{kind=link}
Peripheral membrane proteins
Examples of proteins that use the major structural features:
Polar pocket
PKC- protrudes into headgroup (not into hydrophobic region)
Hydrophobic protrusion
Epsins- ENTH domain protrudes into headgroup and slightly in hydrophobic region
Basic residues binding anionic PL head
Amphiphysin- BAR domain sits on surface and binds via charge interactions
Slide 32
Lipid binding domains of proteins
{kind=link}
Plekstrin- PLC binds phosphatydyl inositols membrane- converts PIP2 into DAG and IP3
C2- binds Ca2+ in PKC
Ankyrin repeat (helix-loop-helix) binds phosphatidylserine, links membrane proteins to cytoskeleton
Grouped into 3 categories:
Signalling (top)
Structural (middle)
Membrane trafficking (bottom)
Slide 33
Lipid binding domain of proteins
Association of protein with membrane is dynamic and depends on:
Type of membrane (what phosphoinositide barcode it has)
[Ca2+]
Availability of lipid species (eg DAG and PIPs)-
Shape of membrane
Proteins don't always bind- bind in opportunistic way and their function is associated with the binding and release of the membrane
Slide 34
2. Proteins lipid-anchored in membrane
Often find more than one lipidation in a protein (many are reversible)
Some lipid anchors are tucked into the protein itself or a chaperone to be freely soluble in cytosol
Looser types of lipid anchors:
Non-permanent binding to membrane, lipid is attached reversibly to protein
Examples of proteins: small/trimeric G-proteins, GPCR C-terminal domains, SNAREs
Examples of anchors:
Palmitoyl group- palmitoylation of any cys in proteins, removed by palmitoyl thioesterase
N-myristoyl group- permanent addition of myristol to alpha-amino group of N-terminal glycine but myristoyl is so short that it isn't hydrophobic enough to permanently stay in membrane
Farnesyl/geranylgeranyl- prenylation of cys of C-X-X-Y at C-termini (farnesylation if Y = A/M/S, geranylgeranyl if Y=L) XXY then excised and methylated, often to small G proteins
All anchors above are on cytosolic side of membrane
Typically found all over the cell (ER, Golgi, PM)
Slide 35
Proteins lipid anchored in membrane
A more permanent lipid anchor is a GPI anchor (Glycosyl phosphatidylinositol)
Example protein is variant surface glycoprotein (VSG) in trypanosoma
Anchors on outside leaflet of membrane (not cytosolic)
GPI-anchors move proteins into lipid rafts
Typically found in PM (not all over cell)
Slide 36
3. Integral membrane proteins (TM)
Most common way of permanently interacting with membrane - TM domain interacts with hydrophobic bilayer
TM proteins have an alpha helical hydrophobic structure- need enough consecutive hydrophobic residues to form helix
Integral proteins are amphipathic- peptide backbone is H bonded tucked inside helix and hydrophobic aa side chains protrude out into bilayer
4 categories of TM protein:
Type I: C- cytosolic side, N-luminal/extracellular (extracellular of cell=inside of ER/Golgi), signal peptide guides N- across membrane into lumen, 1 TM domain
Type II: N- cytosolic, C-luminal, signal peptide guides to membrane but stays on cytosolic side
Type III: TM helix starts at N- (leave just enough to guide protein into membrane) , tail-anchored proteins is when helix starts at C- (helix is last thing synthesised and no classic peptide signal)
Type IV: More than 1 TM domain, C- and N- can be on either side
Slide 37
Integral membrane proteins
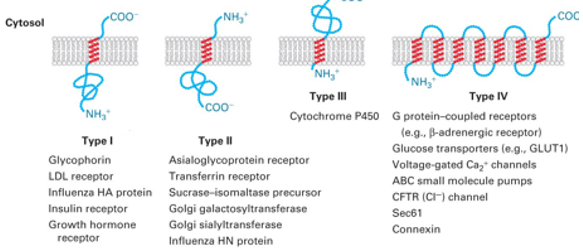{kind=link}
Slide 38
Integral membrane proteins
Features of a TM alpha helix-~20 hydrophobic aa
How to find TM domain:
Hydrophobicity plot used to calculate hydrophobicity of ~20 residues: identify stretches with ΔG >~85 kJ/mol (E required to move stretch out of membrane into water)
Eg GPCR hydropathy plot shows 7 peaks= 7 TM helices
Length can vary of TM domains: PM is thicker than ER- needs longer helices, TMs can be tilted-helices will be longer than thickness of membrane
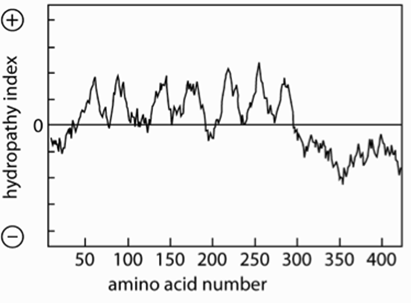{kind=link}
Slide 39
Integral membrane proteins
Other features of TM alpha helix:
Positive inside rule- net charge difference between aa on the cytosolic and outside ends of the helix, helix is more positive on the inside
Aromatic aa often have lipid head groups
Non-hydrophobic aa in membrane helices have functional roles- eg charged residues, +ve charges in voltage sensor of some channels
Slide 40
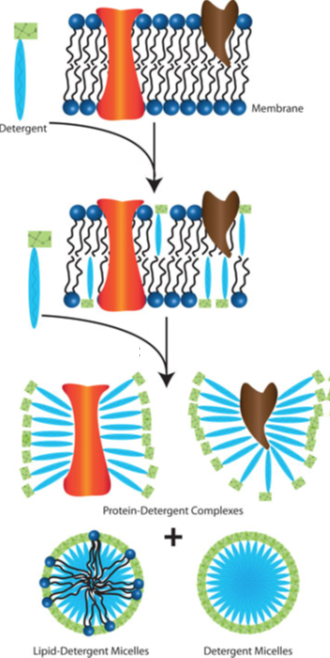{kind=link}
Solubilising TM proteins
To remove TM helix from membrane, need to disrupt membrane (solubilisation) using detergent eg SDS:
Low amount of detergent to membrane= inserts into membrane
Medium amount of detergent= detergent-doped membrane breaks up and get mixed micelles (lipid, detergent and protein)
High amount of detergent= excess detergent removes all lipids into mixed micelles (lipid/detergent) and detergent covers hydrophobic TM domain of proteins
Slide 41
Lipid rafts
Solubilisation allows the purification of TM proteins
But some parts of membranes resist solubilisation in some detergents (Triton-X-100) Detergent Resistant Membranes (DRMs) or Lipid Rafts
Mainly eukaryotic proteins resist solubilisation- lipid rafts are a specific feature of the PM
Slide 42
Lipid rafts: Organisation
Strong association between some lipids breaks membrane fluidity and generates rafts, consist of:
Sphingomyelin lipid (+ve head that can be modified with carbohydrates)
Rich in cholesterol= cholesterol OH H-bonds with amino group of sphingomyelin- keeps rigid (fluid-mosaic model breaks down)
Protein binds lipids via protein TM domain(motifs for cholesterol or lipid) or GPI anchors
Glycolipids (on outside) and actin cytoskeleton (on inside) modulate/shape the raft and recruit proteins
Slide 43
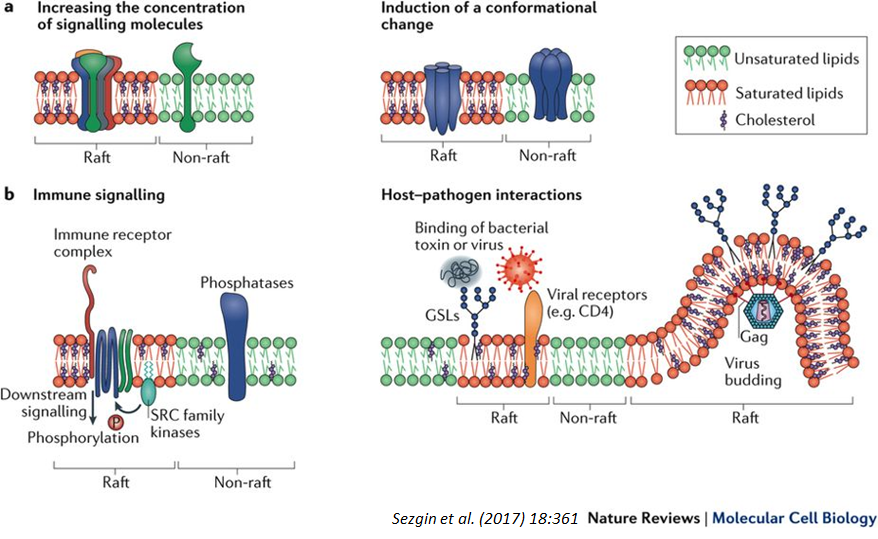
Lipid raft: Functions
Signalling:
Platform for assembling signalling proteins
Increase efficiency by increasing conc of signalling molecules (bring them close together)
Immune signalling
Protein function:
Rigidity alters protein conformation
Host/pathogen interactions:
Ideal structured environment for pathogen binding and virus budding
Eg cholera toxin binds glycolipids (often found in rafts)
Slide 44
Lipid raft: Functions
{kind=link}
Slide 45
Transport across membranes
Membranes separate inside of the cell from the outside, need transport across:
Water soluble molecules/ions= need proteins
Water= need protein channels
Transport proteins can actively transport to increase the concentration
Organelles= have transporter proteins for specific molecules (eg nucleotide-sugar donors in Golgi)
Lipids need special transporters to generate lipid asymmetry
Selective membrane transport can generate gradients, including ionic gradients needed for membrane potential
Slide 46
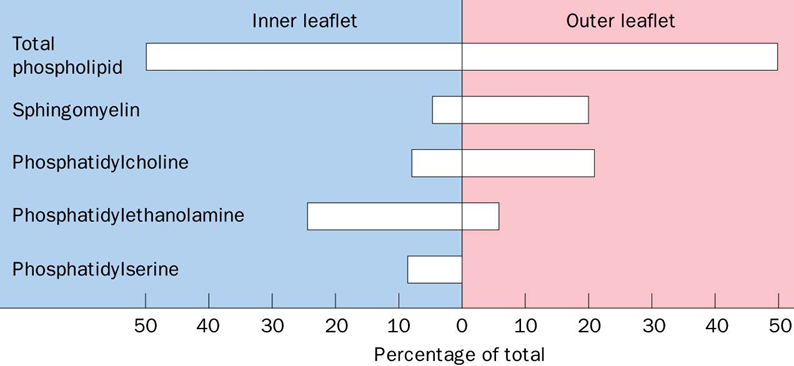
Lipid asymmetry
Synthesis of many PLs occur on cytoplasmic face of ER(outer leaflet of ER), so PE and PS are on correct leaflet of PM(inner PM)
Sphingolipids are synthesised on lumenal side(inside) of ER/Golgi membranes
Cytoplasmic (inner leaflet) of PM often contains more -ve charged amino-phospholipids (eg PS)
PS not on inner leaflet of PM marks a dying cell
{kind=link}
Slide 47
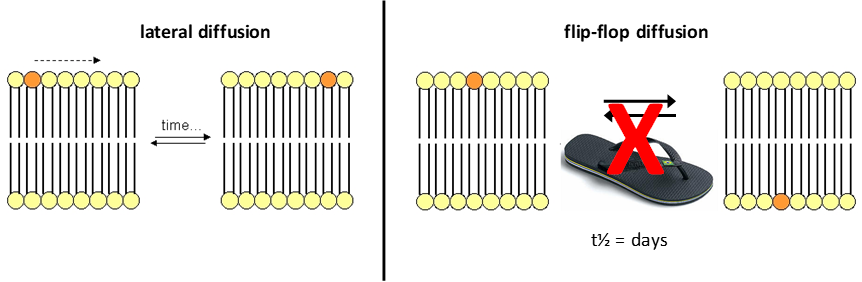
Generating PL asymmetry
Lateral diffusion- free diffusion of PLs laterally in fluid membrane (seconds/minutes)
Flip-flop diffusion- requires transport (days when spontaneous)
Flip-flop diffusion transport:
Scramblases- equilibrate asymmetry of PLs across bilayer
Active transport (flippases and floppases)- use ATP hydrolysis to transport specific PLs across bilayer
{kind=link}
Slide 48
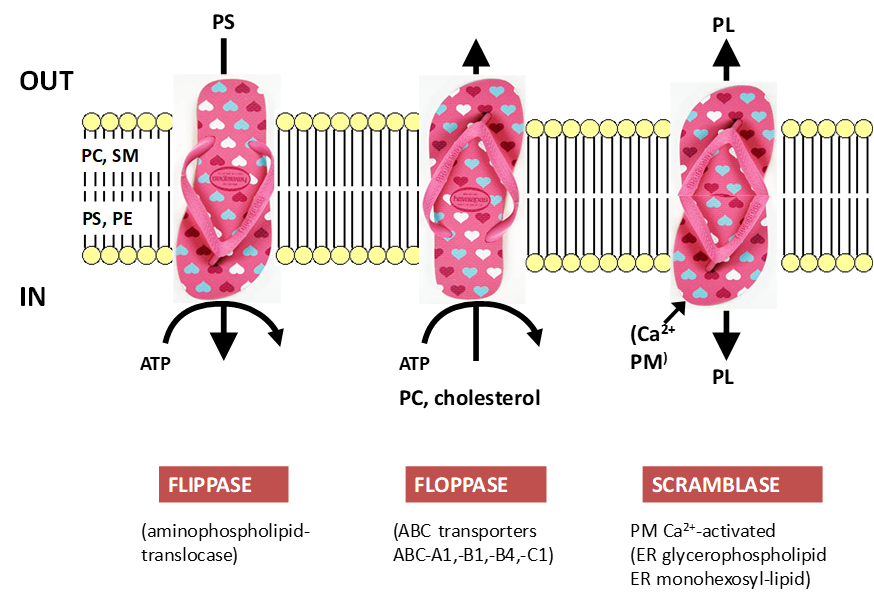
Generating PL asymmetry
{kind=link}
Flippases:
Transport from out to in ( 1 ATP= 1PL)
Eg keeps PS on inner leaflet
Most selective lipid transporters
Floppases:
Transport from in to out (Using ATP)
Eg ABC-B4 moves cholesterol to outside
Scramblases:
No energy and Ca2+ activated
Eg transport PLs from synthesis in ER
Transport from one place to another, don't need to maintain symmetry
Slide 49
Transport of other solutes
1. Pumps(active)
Normally use ATP in intracellular and plasma membranes
Generate electrochemical potential differences for ions and other solutes (gradients): H+ pumping ATPase, K:Na ATPase, ABC transporters
Typically high affinity- so often slow
Operate in one direction (except K:Na ATPase)
Slide 50
Transport of other solutes
2. Carriers (for solute, S)
Intermediate to high affinity
Not (directly) energised
1. Uniport/ facilitated diffusion (passive) = transporting one type of solute (eg Glut4 for glucose uptake)
Energised by electrochemical potentials for ions= one has gradient that helps to transport other:
2. Antiport/Counter-transport (active) eg sugar nucleotide vs free nucleotide in the Golgi
3. Symport/Co-transport (active)
Caption: :
Caption: :
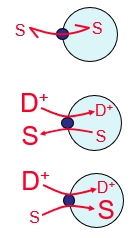{kind=link}
Slide 51
Transport for other solutes
3. Channels (for ions and water)
Transport is thermodynamically dissipative (downhill)- not directly energised but relies on gradient of solute
Open/shut kinetics (gating)
Specificity for ions
Low affinity- transient binding(fast turnover), need some affinity to ensure specificity
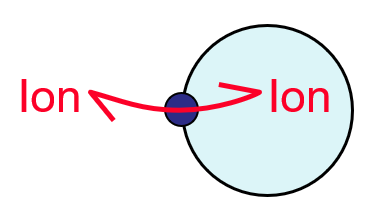{kind=link}
Slide 52
Turnover rate and protein density
Primary pumps:
Turnover rate= slowish enzyme 100/s
Highest density
Carriers:
Turnover rate= little faster 1000/s
Channels:
Turnover rate= catalyse rapid ion movement (approaching diffusion limit), 10^6-10^8 ions/s, 10^9 water/s
Lowest density
Slide 53
ABC transporters (pump family)
Ubiquitous: a diverse class, a superfamily, presence of ATP Binding Cassette in primary structure GX(S,T)GXGK(S,T)(S,T)
Large range of substrates and functions
Eg Used for generating drug resistance (P-glycoprotein)= pump cancer drugs out of cell
{kind=link}
Slide 54
ABC transporters: Cycle
ATP binding = conformational change, alters mem chamber access and affinity
MDR protein:
Picks up substrate on cytosolic side and moves it to cell exterior (after ATP hydrolysis)
ATP binding pulls NBDs together (nucleotide binding domain)- channel shuts on inside
ATP hydrolysis flips pore from internal to external access (changing affinity)- channel opens on outside
Pi release opens NBDs again and next cycle- channel shuts on outside and opens on inside
Slide 55
ABC transporters: Cycle
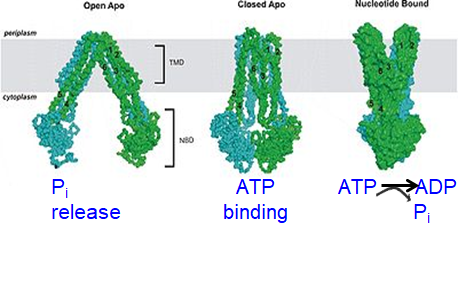{kind=link}
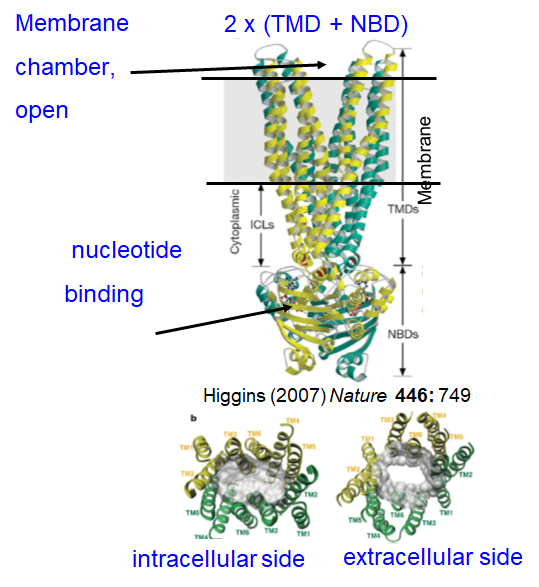{kind=link}
Slide 56
Water transport
Why is osmosis rapid when membranes have hydrophobic interior?
H-bonding properties of water allow solubility in hydrophobic layer
Presence of specific water channels- aquaporins
Discovery of aquaporins:
Experiment with frog oocytes in hypoosmotic solution
Oocyte with aquaporin expands and oocyte without doesn't swell
Features of aquaporins:
Pore selectivity filter ~0.3nm (H2O ~0.28nm)- high selectivity for water because channel has same diameter as water
Low H+ permeability- due to channel size and pore is lined with hydrophobic residues so little interaction for H+
Very high turnover (~10^9)
Slide 57
Summary
Describe the structural features of different lipids and appreciate how these features enable the formation and variability of biological membranes
Cholesterol, sphingolipids and glycolipids form lipid rafts.
Understand the significance of the fluid mosaic model of biological membranes – how it explains the features necessary for a functioning biological membrane and what its limitations are
Lipid rafts a stable membrane domains where membrane fluidity is restricted.
Evaluate the molecular features and energetic barriers for lipid transport between and across membranes
Lipid asymmetry is generated using flippases and floppases and uses ATP.
Appreciate how proteins interact with biological membranes
Integral membrane proteins have helical transmembrane domains of ~20 amino acids. Peripheral membrane proteins bind lipids. Proteins can have covalently linked lipid anchors or GPI anchors.
Understand how different classes of molecules cross biological membranes
Hydrophobic molecules can diffuse through the bilayer. Ions and hydophilic molecules use transport protein. Channels are fast, selective and passive. Carriers are slower, and use one gradient to carry a second solute. Pumps are slow, but use ATP and can achieve high accumulation. Water needs a channel to cross membranes.
Want to create your own Slides for free with GoConqr? Learn more.