6480547
Question 1
Question
Most types of molecule and cellular component of animal cells are affected during [blank_start]ageing[blank_end], and different types of animal cells have [blank_start]different lifespans[blank_end].
Answer
-
ageing
-
different lifespans
Question 2
Question
Cellular ageing is studied using cells from model organisms that have very different lifespans.
Answer
- True
- False
Question 3
Question
Two of the many theories of ageing are the n[blank_start]etwork theory[blank_end], which states that cellular ageing is the result of a [blank_start]combination of processes[blank_end], and the f[blank_start]ree radical theory[blank_end], according to which [blank_start]free radical damage[blank_end] is the cause of [blank_start]molecular and cellular ageing[blank_end].
Answer
-
etwork theory
-
combination of processes
-
ree radical theory
-
free radical damage
-
molecular and cellular ageing
Question 4
Question
Genes that confer protection against [blank_start]free radicals[blank_end] and other stresses may have a positive effect against molecular and cellular damage occurring during [blank_start]ageing[blank_end], but will only be selected for by evolution if they also promote [blank_start]reproductory fitness[blank_end].
Answer
-
free radicals
-
ageing
-
reproductory fitness
Question 5
Question
Vertebrate tissues have a mixture of [blank_start]post-mitotic[blank_end] non-dividing cells, such as [blank_start]neurons[blank_end] and skeletal muscle cells, and [blank_start]dividing[blank_end] cells, such as [blank_start]epithelial cells[blank_end] (and germ cells).
Answer
-
post-mitotic
-
neurons
-
epithelial cells
-
dividing
Question 6
Question
Label the diagram of formation of Reactive Oxygen Species during oxidative phosphorylation at the mitochondria
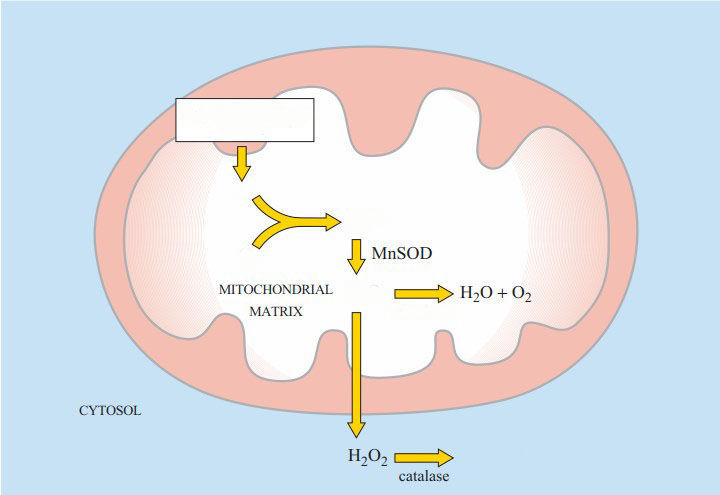{kind=link}
Answer
-
electron transport chain
-
e-
-
O2
-
O2•-
-
H2O2
-
catalase
-
H2O + O2
-
superoxide dismututase
Question 7
Question
Free [blank_start]radicals[blank_end] are atoms or molecules that possess [blank_start]unpaired[blank_end] electrons and are therefore very [blank_start]reactive[blank_end].
Answer
-
radicals
-
unpaired
-
reactive
-
unreactive
Question 8
Question
There are many naturally occurring [blank_start]free radicals[blank_end]; one major source within cells is [blank_start]oxidative phosphorylation[blank_end], which takes place in the inner [blank_start]mitochondrial[blank_end] membrane.
Answer
-
free radicals
-
oxidative phosphorylation
-
mitochondrial
-
cell
Question 9
Question
Free radicals cause [blank_start]oxidative[blank_end] damage to all types of molecules and cell organelles.
Particularly: DNA damage - [blank_start]double strand breaks[blank_end] and altered bases/nucleosides, chain reactions of free radical formation in [blank_start]lipid peroxidation[blank_end], damage to amino acids causing changes to [blank_start]activity/conformation[blank_end] of the protein.
Answer
-
oxidative
-
double strand breaks
-
lipid peroxidation
-
activity/conformation
Question 10
Question
Cellular defences against free radicals have evolved; these are either primary or secondary defences. Primary defences include small-molecule radical [blank_start]scavengers[blank_end], proteins that bind [blank_start]metal ions[blank_end], and [blank_start]antioxidant enzymes[blank_end]. Secondary defences include [blank_start]repair enzymes[blank_end], [blank_start]stress response[blank_end] proteins and [blank_start]protein degradation[blank_end] systems.
Answer
-
scavengers
-
metal ions
-
antioxidant enzymes
-
repair enzymes
-
stress response
-
protein degradation
Question 11
Question
[blank_start]Replicative senescence[blank_end] is the irreversible state reached by proliferative cells when they withdraw from the [blank_start]cell cycle[blank_end], and do not undergo any further divisions. It has been studied by measuring [blank_start]population doublings[blank_end] of cells in culture.
Answer
-
Replicative senescence
-
cell cycle
-
population doublings
Question 12
Question
Senescent cells exhibit changes in [blank_start]gene expression[blank_end], which may not only affect their function, but may also affect [blank_start]surrounding cells[blank_end]. They also look different from [blank_start]dividing cells[blank_end], being bigger and having larger nuclei.
Answer
-
gene expression
-
surrounding cells
-
dividing cells
Question 13
Question
Several different events can cause cells to become [blank_start]senescent[blank_end]; these include [blank_start]telomere[blank_end] shortening, some types of DNA [blank_start]damage[blank_end], decondensation of [blank_start]chromatin[blank_end], overactivity of some [blank_start]mitogenic stimuli[blank_end] and activation of some [blank_start]oncogenes[blank_end].
Answer
-
senescent
-
telomere
-
damage
-
chromatin
-
mitogenic stimuli
-
oncogenes
Question 14
Question
[blank_start]Telomeres[blank_end] may be preferentially susceptible to [blank_start]DNA damage[blank_end], and therefore not only trigger [blank_start]replicative senescence[blank_end] due to [blank_start]shortening[blank_end] caused by repeated cell division, but also act as a type of ‘sensor’ of DNA damaging events, such as [blank_start]oxidative stress[blank_end].
Answer
-
Telomeres
-
DNA damage
-
replicative senescence
-
shortening
-
oxidative stress
Question 15
Question
[blank_start]Post-mitotic[blank_end] cells such as [blank_start]neurons[blank_end] and skeletal muscle cells exhibit a number of changes during ageing. These changes include [blank_start]mitochondrial damage[blank_end], abnormalities in [blank_start]protein folding[blank_end], protein [blank_start]accumulation[blank_end], and [blank_start]protein[blank_end] glycation.
Answer
-
Post-mitotic
-
neurons
-
mitochondrial damage
-
protein folding
-
accumulation
-
protein
Question 16
Question
[blank_start]Mitochondrial[blank_end] function is impaired with age, and genes encoded by [blank_start]mtDNA[blank_end] are particularly vulnerable to [blank_start]oxidative damage[blank_end].
Answer
-
Mitochondrial
-
mtDNA
-
oxidative damage
Question 17
Question
Changes in protein [blank_start]folding[blank_end] and turnover occur with increasing age. perhaps due to problems with [blank_start]chaperones[blank_end] or the protein [blank_start]degradation[blank_end] system. Accumulation of misfolded or [blank_start]damaged[blank_end] proteins therefore often occurs in long-lived cells such as [blank_start]neurons[blank_end]. These can form insoluble [blank_start]aggregates[blank_end] called [blank_start]amyloid[blank_end] fibrils or plaques, causing [blank_start]disease[blank_end].
Answer
-
folding
-
chaperones
-
degradation
-
damaged
-
neurons
-
aggregates
-
amyloid
-
disease
Question 18
Question
Interaction of [blank_start]sugars[blank_end] with amino acid residues starts a series of reactions leading to [blank_start]protein glycation[blank_end] and the accumulation of [blank_start]AGEs[blank_end] (Advanced Glycosylation End-products). Long-lived proteins (e.g. [blank_start]extracellular matrix proteins[blank_end]) are particularly susceptible to damage by [blank_start]glycation[blank_end].
Answer
-
sugars
-
protein glycation
-
AGEs
-
extracellular matrix proteins
-
glycation
Question 19
Question
Both mitotic and post-mitotic cells can be affected by genomic instability.
Answer
- True
- False
Question 20
Question
Many segmental [blank_start]progeroid[blank_end] syndromes in humans are due to [blank_start]mutations[blank_end] in genes encoding proteins that play a role in the detection of [blank_start]DNA damage[blank_end], or DNA [blank_start]repair[blank_end]. This suggests that DNA damage plays a role in [blank_start]normal ageing[blank_end].
Answer
-
progeroid
-
mutations
-
DNA damage
-
repair
-
normal ageing
Question 21
Question
A [blank_start]mutation[blank_end] in the gene encoding nuclear A-type lamins has also been found in one [blank_start]progeroid[blank_end] syndrome, [blank_start]HGPS[blank_end]. [blank_start]Nuclear lamins[blank_end] are [blank_start]intermediate filaments proteins[blank_end], which form a structural lattice attached to the inner face of the [blank_start]nuclear envelope[blank_end] and play a role in cell [blank_start]division[blank_end] and gene [blank_start]expression[blank_end].
Answer
-
mutation
-
progeroid
-
HGPS
-
Nuclear lamins
-
intermediate filaments proteins
-
nuclear envelope
-
division
-
expression
Question 22
Question
The [blank_start]insulin/IGF-1-like[blank_end] signalling pathway in C. elegans , D. melanogaster and mice affects both [blank_start]stress responses[blank_end] and longevity.
Activation of this pathway in these three laboratory model organisms leads to [blank_start]downregulation[blank_end] of stress response genes and [blank_start]reduces[blank_end] lifespan.
Although an [blank_start]analogous[blank_end] pathway occurs in humans, and affects [blank_start]growth[blank_end] and metabolism, the effects on stress response genes and [blank_start]longevity[blank_end] are not known.
Answer
-
insulin/IGF-1-like
-
stress responses
-
downregulation
-
reduces
-
analogous
-
growth
-
longevity
Question 23
Question
mtDNA is more vulnerable to oxidative damage because:
Answer
-
mtDNA is not complexed with histones, which in nuclear chromatin, offer some protection against oxidative damage
-
almost all mtDNA is coding DNA, so any mutations that occur may affect gene products
-
DNA repair is not as efficient in the mitochondria, as not all DNA repair mechanisms take place
-
mtDNA is just more delicate than nuclear DNA
Question 24
Question
[blank_start]mtDNA[blank_end] codes for several [blank_start]proteins[blank_end] that are required in the [blank_start]TCA[blank_end] cycle and the [blank_start]electron transport[blank_end] chain. So if mtDNA is damaged by [blank_start]ROS[blank_end], this has huge repercussions on the cell's ability to [blank_start]generate energy[blank_end].
Combined with other age related damage to [blank_start]mitochondria[blank_end] in [blank_start]post-mitotic[blank_end] cells, to the membrane and proteins, this can be a real problem.
Answer
-
mtDNA
-
proteins
-
TCA
-
electron transport
-
ROS
-
generate energy
-
mitochondria
-
post-mitotic
Want to create your own Quizzes for free with GoConqr? Learn more.